Life Extension Magazine®
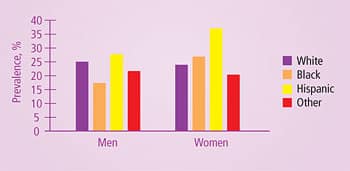
| Can you control your genetic destiny? Are you doomed to suffer debilitating diseases because of “bad genes”? The answer is that a powerful strategy called phenotypic nutrition can help modulate the expression of your unique genetic code, thus dramatically reducing your risk of developing disease. Phenotypic nutrition uses specific nutrients with targeted biochemical and genetic effects to help protect you from disease. One of the most important threats to your health and longevity is metabolic syndrome, a disorder little known to the public but fast becoming an extremely important public health issue. This deadly, common condition affects approximately one of every five people overall, with even higher rates among certain ethnic groups.1 By applying a phenotypic nutrition strategy, we can help to guard against the deadly dangers of cardiovascular disease associated with metabolic syndrome. Metabolic Syndrome DefinedOver 20 years ago, Stanford University professor Gerald Reaven, MD, coined the term “Syndrome X” to describe a constellation of metabolic abnormalities in serum cholesterol, blood pressure, blood-clotting tendencies, and cardiovascular disease risk.2 Now known as metabolic syndrome, this constellation of abnormalities is recognized as placing individuals at dramatically in-creased risk for cardiovascular disease.3 For example, a study published in 2004 in the Journal of the American Medical Association followed more than 1,200 men for 11 years. The investigators found that men with metabolic syndrome were 160-320% more likely to die from coronary heart disease than those without metabolic syndrome.4 The diagnostic criteria for metabolic syndrome proposed by the Third Report of the National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults (Adult Treatment Panel III) include:
Despite the grave cardiovascular risk posed by metabolic syndrome, there is currently no standard, accepted interventional treatment regimen to prevent the disorder. However, for those at greatest risk for metabolic syndrome, phenotypic nutrition allows us to tailor a nutrient and supplement strategy to amplify beneficial biochemical pathways as well as gene expression.
Orthomolecular Medicine: Visionary ScienceIn 1968, the Nobel Prize-winning scientist Linus Pauling, PhD, published an article in the journal Science describing the “orthomolecular” approach to illness. This was the first time that such an approach was presented to the public—specifically, that “varying the concentrations of substances normally present in the human body may control mental disease.”6 Dr. Pauling later extended this strategy beyond mental illness to include the discipline of orthomolecular medicine. Orthomolecular medicine is a paradigm that attempts to prevent and treat disease by integrating conventional medical therapies with vitamins, phytonutrients, and other dietary micronutrients. Dr. Pauling’s fascination with vitamin C and its role in helping to prevent and treat disease is an example of orthomolecular medicine. Dr. Pauling published pioneering reviews on the use of vitamin C and the orthomolecular approach to cancer over 30 years ago.7,8 Biochemical Variations and Nutrient NeedsFrom our physical and psychological traits to our enzyme systems that process nutrients and help guard against disease, each of us is unique. Roger Williams, PhD, discoverer of pantothenic acid (vitamin B5) and founder and director of the Clayton Foundation Biochemical Institute at the University of Texas, was a visionary in the understanding of biochemical uniqueness. Dr. Williams’ timeless classic Biochemical Individuality, originally published in 1956, acknowledges the unique biochemical variations among individuals and how these differences relate to their distinct nutritional needs for optimal health and disease prevention.
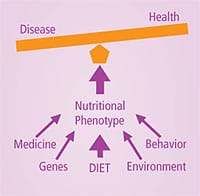
Nutrigenomics: Guiding Genetic DestinyDriven by recent technological breakthroughs associated with the mapping of the human genome, the science of nutrigenomics holds great potential for predicting how specific nutrients and dietary ingredients can directly affect health and disease by specific genetic interactions. Nutrigenomics holds promise in advancing the goal of preventing disease with individualized, targeted nutrient-gene interactions.9,10 Nutrigenomics suggests that genetic testing will allow for “individualized nutrition” based on unique genetic needs. Currently, however, nutrigenomics remains a very young and developing science that has not yet developed to the point of being able to offer broad-based nutrient-gene testing of this sort. Furthermore, ethical questions surround genetic testing. For example, if you were to undergo a genetic test predicting you were at fourfold greater risk of having a heart attack unless you consumed specific nutrients, would you want that information shared with your health insurance company or employer? Who would control the confidentiality of this type of information? There is, however, another approach that we can use for specific, individualized nutrient recommendations that incorporates Dr. Pauling’s visionary approach to orthomolecular medicine, Dr. Williams’ brilliant insight into biochemical individuality, and the futurist promise of nutrigenomics—without having to worry about the ethical and confidentiality issues related to genetic testing. This alternative approach is called phenotypic nutrition. Genes, Genotype, and PhenotypeYour unique genetic code defines your genotype. The expression of your individual genetic code is your phenotype. Both your genes and your environment influence your phenotype. While it is not known exactly which genes are responsible for the constellation of abnormalities associated with metabolic syndrome, a number of genes so far identified play a role in the malfunction associated with abnormally elevated plasma glucose.11 Metabolic syndrome is a phenotypic expression of the genetic code as it interacts with the environment. Specifically, if you have a genetic tendency to develop the metabolic syndrome phenotype, you will be far more likely to develop this phenotype if you are overweight, do not exercise, consume a diet high in simple sugars and saturated fats, and do not consume enough specific nutrients with beneficial nutrient-gene interactions. In 2004, an innovative approach to weight management and obesity was published in the peer-reviewed journal Current Drug Targets. This approach, called “nutriphenotypic,” is selective nutrition based on an individual’s phenotypic metabolic characteristics.12 This past year, researchers described the concept of a “nutritional phenotype” that characterizes the relationship of biochemical measures, metabolic parameters, and functional characteristics (for example, exercise, body weight, exposure to environmental pollutants and toxins, emotional stress) on health. The nutrient phenotype is a function of genes, diet, environment, and behavior.13
The nutriphenotypic/nutrient phenotype approach represents an integrative strategy to help prevent and treat disease. Just as specific inputs to a computer will cause specific outputs, phenotypic nutrition uses specific nutrients for specific effects. Phenotypic nutrition affects the expression of your genes and unique biochemistry so that you can influence your genetic destiny. Instead of dooming individuals to disease and an early demise due to “bad genes,” phenotypic nutrition uses nutrients that act on specific genes as well as biochemical and molecular pathways to help prevent disease and achieve optimal health and longevity. With phenotypic nutrition, once high-risk individuals have been identified, strategies that include dietary modifications and specific nutraceuticals can be implemented to help decrease disease risk. Identifying Those Most at RiskMetabolic syndrome is characterized by insulin resistance. Identifying individuals with evidence of insulin resistance before the development of full-blown metabolic syndrome allows us to intervene with a nutrient and lifestyle strategy to prevent the progression of this condition. Thankfully, simple metabolic markers are very useful in helping to identify individuals with insulin resistance who are at high risk of cardiovascular disease.
Moreover, research has demonstrated that a condition of low-grade systemic inflammation is associated with insulin resistance. For example, a recent clinical study showed that insulin and insulin resistance remained significantly and independently related to C-reactive protein (CRP) levels, a marker of inflammation, after adjustments for age, sex, body mass index, waist size, alcohol consumption, level of physical activity, and smoking habits.15 Another clinical study found that CRP was significantly correlated to insulin resistance.16
DHEA. Scientists have associated suboptimal levels of dehydroepiandrosterone (DHEA) with risk for metabolic syndrome. For example, a cross-sectional study of 400 independently living men aged 40-80 reported a statistically significant relationship between DHEA levels and risk of metabolic syndrome—specifically, the higher the level of DHEA, the lower the risk of metabolic syndrome.17 A number of randomized, placebo-controlled clinical studies have shown that supplemental DHEA excels in improving many metabolic derangements associated with metabolic syndrome. For example, in a study of elderly men and women, 50 mg per day of DHEA for six months was associated with significant improvements in visceral fat, subcutaneous fat, and insulin sensitivity.18 Another study evaluated DHEA’s effects on markers of insulin resistance and cardiovascular risk in 28 hypoadrenal (demonstrating diminished adrenal gland function) women. In this randomized, double-blind, placebo-controlled crossover study, women supplementing with DHEA significantly improved their insulin sensitivity and reduced their levels of triglycerides and low-density lipoprotein (LDL).19 These simple, surrogate screening assessments for insulin resistance and cardiovascular disease risk—that is, of triglycerides, insulin, HDL, CRP, and DHEA—can identify those at high risk of developing metabolic syndrome. DHA and EPAPeroxisome proliferator-activated receptors (PPARs) are genetic transcription factors that act on DNA response elements. Carbohydrate responsive element binding protein (ChREBP) and sterol regulatory element binding protein-1c (SREBP-1c) are two other critical regulators of glucose metabolism in the liver. Fatty acids such as DHA (docosahexaenoic acid) and EPA (eicosapentaenoic acid) are nutrients that directly affect these genetic transcription factors. PPARs are present in high-energy-demand tissues, including the liver, heart, and, to a lesser extent, skeletal muscle. PPARs are critical to a phenotypic nutrition approach to preventing metabolic syndrome, as PPARs promote fatty acid oxidation and fat cell differentiation while promoting glucose utilization.20 Studies have shown that EPA and DHA decrease the expression of genes involved in fatty acid storage. Through a depleting effect on SREBP-1c, EPA and DHA reduce insulin response to oral glucose and protect against the effect of insulin resistance induced by a high-fat diet. At the molecular level, this is accomplished by preventing the decreased expression of the gene encoding the glucose transporter GLUT4 in fat cells.21 Research has also shown that EPA and DHA down-regulate genes involved in inflammation, an important protective attribute for our prevention program, given inflammation’s association with metabolic syndrome. For example, pre-clinical studies have shown that these fatty acids down-regulate pro-inflammatory nuclear factor-kappa beta (NfkB) through a PPAR-gamma-dependent pathway.22 Human studies have shown a relationship between increased DHA consumption and diminished CRP levels,23 and other in-vivo studies have shown that markers of oxidative stress such as urinary F2-isoprostanes decrease significantly with EPA and DHA intake.24
Lipoic AcidA naturally occurring nutrient, lipoic acid is known to improve glucose metabolism by influencing genetic transcriptional factors in fat cells through the mitogen-activated protein kinase (MAPK) pathway.25 For example, in a multicenter, placebo-controlled trial, type II diabetes patients treated with lipoic acid demonstrated a significant increase in insulin-stimulated glucose disposal.26 Furthermore, lipoic acid’s molecular attributes include increasing glucose uptake through recruitment of the glucose transporter-4 (GLUT-4) to plasma membranes, a mechanism that is shared with insulin-stimulated glucose uptake.27 Research also shows that lipoic acid decreases markers of vascular inflammation in metabolic syndrome. A recent randomized, double-blind clinical trial showed that after four weeks of therapy, endothelium-dependent vasodilation of the brachial artery was increased by 67%, 44%, and 75% in groups receiving irbesartan (an angiotensin-blocking drug), lipoic acid, and irbesartan plus lipoic acid, respectively, compared to placebo. Furthermore, treatment with irbesartan and lipoic acid was associated with statistically significant reductions in plasma levels of pro-inflammatory mediators such as interleukin-6.28 | |||||||



Glucose and Dietary CarbohydratesThe Mediterranean diet is rich in whole grains, fruits, vegetables, and fat from fish, nuts, and olives. Low in rapidly digested simple carbohydrates, this diet is helpful in decreasing genetic expression of fat-storage enzymes activated by high amounts of dietary carbohydrates. Scientists have realized that in addition to the key regulatory hormone insulin, glucose itself plays a critical role in helping to regulate glucose metabolism and fat storage (lipogenesis). Decreased ChREBP gene expression results in decreased fat-storage gene expression, thereby demonstrating the direct implication of ChREBP in glucose action and fat storage.29 Randomized clinical trials have shown that high-carbohydrate/low-fat diets accentuate the metabolic abnormalities of patients with glucose dysregulation such as diabetes. For example, a trial evaluated patients with type II diabetes mellitus after they were randomly placed on diets comprising either 55% carbohydrates, 30% fat, and 15% protein, or 40% carbohydrates, 45% fat, and 15% protein for six weeks, followed by crossover to the other diet. The high-carbohydrate/low-fat diet in patients with type II diabetes led to:
Simply stated, in people with impaired blood glucose control, a diet high in carbohydrates will cause gene expression to activate enzymes involved in fat storage. Furthermore, a diet moderate in carbohydrates and relatively high in monounsaturated fat from olive oil and polyunsaturated fatty acids from fish, such as the Mediterranean diet, has been shown in prospective, randomized clinical trials to decrease insulin resistance, improve endothelial function, and decrease markers of inflammation.31 Also, this dietary approach may be better than the National Cholesterol Education Program’s Step 1 prudent diet in preventing cardiac events.32
Other prospective clinical trials have shown that replacing carbohydrate calories with calories from nuts such as almonds, which are rich in monounsaturated and polyunsaturated fats, reduces many of the metabolic abnormalities associated with metabolic syndrome.33 Clearly, a Mediterranean-type diet low in simple sugars and rich in vegetables, fruits, fish, nuts, olive oil, and whole grains is an ideal dietary strategy for helping to prevent metabolic syndrome. BioflavonoidsInflammation and associated insulin resistance play a critical role in the development of metabolic syndrome and type II diabetes.34,35 Nutrients that act to down-regulate genes involved in inflammation are critical to preventing metabolic syndrome. Quercetin, a potent bioflavonoid found in vegetables, has been shown to inhibit the genetic expression of pro-inflammatory cytokines through inhibition of the NfkB pathway both in cell studies and animal trials.36 Along with quercetin, the bioflavonoid resveratrol, found in the skin of red fruits like grapes, has been shown in molecular pre-clinical studies to inhibit NfkB and cAMP response element binding protein-dependent, pro-inflammatory gene transcription to a greater extent than the potent cortico-steroid dexamethasone.37 The benefits of olive polyphenols are striking. Olive polyphenols have direct effects on the expression of genes involved in the vascular endothelium (the lining of blood vessel walls). At nutritionally relevant concentrations, hydroxytyrosol, a principal polyphenol in olives, reduced cell “stickiness” to the vascular endothelium by decreasing genetic expression for NfkB of vascular cell adhesion molecule-1 (VCAM-1) messenger RNA (mRNA).38 Furthermore, pre-clinical studies have shown that olive oil with a higher content of polyphenolic compounds shows protective effects in models of inflammation.39 Other pre-clinical studies have shown that olive polyphenols dramatically increase the resistance of LDL to oxidation.40 Oxidized LDL is a potent trigger for atherosclerosis.41
TocopherolsVitamin E is an important nutrient in preventing metabolic syndrome, as it helps regulate several genes affecting cardiovascular disease risk:42
Furthermore, the appropriate genetic regulation of glucose transport protein (GLUT-3) is critical to optimal blood glucose control. Studies have shown that both aging and vitamin E deficiency are associated with decreased expression of GLUT-3.43 Carotenoids and RetinoidsUsing data from the Third National Health and Nutrition Examination Survey (1988-1994), researchers evaluated the intake of vitamins A and C, retinyl esters, five carotenoids, and other trace nutrients in 8,808 US adults aged 20 and older with and without metabolic syndrome. After adjusting for factors like age, sex, ethnicity, education, smoking status, and physical activity, they found that individuals with metabolic syndrome had significantly lower concentrations of retinyl esters, vitamin C, and carotenoids, except lycopene.44 Vitamin A is particularly important in preventing metabolic syndrome, given research showing that vitamin A plays a role in the regulation of genes involved in systemic insulin resistance. Specifically, researchers have shown that retinoic acid, the acid form of vitamin A, is a signal that inhibits the expression of resistin, an adipocyte-secreted protein previously proposed to act as an inhibitor of adipocyte differentiation and as a systemic insulin resistance factor.45 Pre-clinical models of early development have also shown that suboptimal intake of vitamin A decreases insulin-producing cells, and that this reduction can be attributed to a reduced rate of fetal beta-cell replication. This could potentially contribute to impaired glucose tolerance later in adult life.46
Cinnamon Extracts
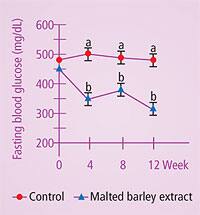
In clinical trials, cinnamon has demonstrated remarkable effects on glucose control. In 2003, a placebo-controlled study of type II diabetes patients who were given one, three, or six grams a day of cinnamon or placebo showed that after 40 days, all three levels of cinnamon reduced mean fasting serum glucose by 18-29%, triglycerides by 23-30%, LDL by 7-27%, and total cholesterol by 12-26%. No significant changes were noted in the placebo groups.47 Cinnamon also has been shown to have excellent antioxidant properties. Natural water-soluble cinnamon extract has been shown to inhibit oxidation by 88%, while a synthetic antioxidant control, butylated hydroxytoluene, inhibited oxidation by only 80%.48 Water-soluble polyphenol polymers (polyphenol type-A polymers) from cinnamon increase insulin-dependent glucose metabolism in vitro. These polymers have recently been characterized by nuclear magnetic resonance and mass spectroscopy.49 The polyphenol type-A polymers from cinnamon up-regulate expression of genes involved in activation (phosphorylation) of the insulin receptor and increase glucose uptake in cell studies.50 In other cell studies, other cinnamon polyphenol polymers such as methylhydroxychalcone have been shown to be potent insulin mimics.51 Barley ExtractsPurine adenine derivatives called cytokinins (not to be confused with the pro-inflammatory glycoproteins called cytokines) have been shown to control ribosomal RNA (rRNA) in barley.52 Furthermore, cytokinin genes in barley have recently been cloned.53 In pre-clinical studies, cytokinin-rich barley extract has been shown to alleviate many diabetes symptoms, including fasting serum glucose, in genetically obese mice, without affecting insulin levels.54 Most compelling is that the barley extract appears to function similarly to the drug metformin, known to dramatically improve plasma glucose in patients with type II diabetes. Cell study data show that barley extract inhibits gluconeogenesis (creation of new glucose molecules) in liver cells.55 Human pilot studies have also been impressive. The preliminary data show that barley extract reduced fasting and postprandial glucose levels in type II diabetes, reduced the LDL/HDL ratio, improved postprandial (after-meal) glucose, and improved glucose transport into muscle cells.55
Coffee PolyphenolsSeveral published studies associate coffee consumption with a dramatically reduced risk of type II diabetes. For example, a large study (involving 16,670 men and women) published in JAMA in 2004 showed a very strong relationship between increased coffee consumption and decreased risk of developing type II diabetes mellitus.56 Another very large study that followed 41,934 men from 1986 to 1998, and 84,276 women from 1980 to 1998, showed a similar, powerful association between greater coffee intake and reduced risk for type II diabetes after adjusting for age, body mass index, and other risk factors.57 Water extracts of roasted coffee residues, including the primary coffee polyphenols caffeic acid and chlorogenic acid, have also been found to scavenge free radicals and offer protection against lipid peroxidation and the oxidative damage of protein.58 Found in relatively abundant amounts in coffee beans, chlorogenic acid is also found in blueberries, apples, and eggplant. Animal studies have shown that chlorogenic acid improves glucose tolerance, decreases fasting plasma cholesterol and triglycerides, and improves mineral pool distribution.59 Clinical studies have shown that chlorogenic acid delays glucose absorption and improves gastrointestinal hormone secretion. For example, a randomized crossover study in healthy volunteers showed that consuming 400 mL of decaffeinated coffee (equivalent to 2.5 mmol of chlorogenic acid per liter) significantly increased glucagon-like peptide 1 (GLP-1) secretion after meals compared to controls.60 The association between chlorogenic acid and GLP-1 is particularly remarkable. Several GLP-1-related pharmaceutical agents are mimicking this hormone as a treatment for type II diabetes mellitus, including the recently FDA-approved GLP-1 analog BYETTA™ (exenatide). Chlorogenic acid acts to increase GLP-1 and delay glucose absorption, thereby maintaining the responsiveness of the insulin-producing beta cells of the pancreas.61 Ferulic acid, another water-soluble coffee extract, improves blood vessel relaxation and decreases levels of homocysteine, an independent cardiovascular risk factor.62 ConclusionYou are not doomed to disease by “bad genes”! Phenotypic nutrition enables you to choose dietary strategies and nutrients that influence powerful biochemical and genetic factors to help control the expression of your genetic code to your benefit. In a prevention program to guard against metabolic syndrome, the first step is identifying if you are at high risk. This can easily be accomplished by checking simple blood tests of triglycerides, insulin, HDL, CRP, and DHEA. Next, select specific nutritional strategies to beneficially influence the expression of your genetic code and biochemical make-up to decrease your disease risk. It is up to you—help control your own genetic destiny, or roll the dice and hope for the best. | ||||||||
| References | ||||||||
| 1. Ford ES, Giles WH, Dietz WH. Prevalence of the metabolic syndrome among US adults: findings from the third National Health and Nutrition Examination Survey. JAMA. 2002 Jan 16;287(3):356-9. 2. Reaven GM. Syndrome X: 6 years later. J Intern Med Suppl. 1994;736:13-22. 3. Eckel RH, Grundy SM, Zimmet PZ. The metabolic syndrome. Lancet. 2005 Apr 16;365(9468):1415-28. 4. Lakka HM, Laaksonen DE, Lakka TA, et al. The metabolic syndrome and total and cardiovascular disease mortality in middle-aged men. JAMA. 2002 Dec 4;288(21):2709-16. 5. Anon. Third Report of the National Cholesterol Education Program (NCEP) Expert Panel on Detection, Evaluation, and Treatment of High Blood Cholesterol in Adults (Adult Treatment Panel III) final report. Circulation. 2002 Dec 17;106(25):3143-421. 6. Pauling L. Orthomolecular psychiatry. Varying the concentrations of substances normally present in the human body may control mental disease. Science. 1968 Apr 19;160(825):265-71. 7. Cameron E, Pauling L. Ascorbic acid and the glycosaminoglycans. An orthomolecular approach to cancer and other diseases. Oncology. 1973;27(2):181-92. 8. Cameron E, Pauling L. The orthomolecular treatment of cancer. I. The role of ascorbic acid in host resistance. Chem Biol Interact. 1974 Oct;9(4):273-83. 9. Fairfield KM, Fletcher RH. Vitamins for chronic disease prevention in adults: scientific review. JAMA. 2002 Jun 19;287(23):3116-26. 10. Muller M, Kersten S. Nutrigenomics: goals and strategies. Nat Rev Genet. 2003 Apr;4(4):315-22. 11. Marx J. Unraveling the causes of diabetes. Science. 2002 Apr 26;296(5568):686-9. 12. Joyal SV. A perspective on the current strategies for the treatment of obesity. Curr Drug Targets CNS Neurol Disord. 2004 Oct;3(5):341-56. 13. Zeisel SH, Freake HC, Bauman DE, et al. The nutritional phenotype in the age of metabolomics. J Nutr. 2005 Jul;135(7):1613-6. 14. McLaughlin T, Abbasi F, Cheal K, et al. Use of metabolic markers to identify overweight individuals who are insulin resistant. Ann Intern Med. 2003 Nov 18;139(10):802-9. 15. Bo S, Gambino R, Uberti B, et al. Does C-reactive protein identify a subclinical metabolic disease in healthy subjects? Eur J Clin Invest. 2005 Apr;35(4):265-70. 16. Temelkova-Kurktschiev T, Siegert G, Bergmann S, et al. Subclinical inflammation is strongly related to insulin resistance but not to impaired insulin secretion in a high risk population for diabetes. Metabolism. 2002 Jun;51(6):743-9. 17. Muller M, Grobbee DE, den T, I, Lamberts SW, van der Schouw YT. Endogenous sex hormones and metabolic syndrome in aging men. J Clin Endocrinol Metab. 2005 May;90(5):2618-23. 18. Villareal DT, Holloszy JO. Effect of DHEA on abdominal fat and insulin action in elderly women and men: a randomized controlled trial. JAMA. 2004 Nov 10;292(18):2243-8. 19. Dhatariya K, Bigelow ML, Nair KS. Effect of dehydroepiandrosterone replacement on insulin sensitivity and lipids in hypoadrenal women. Diabetes. 2005 Mar;54(3):765-9. 20. Ferre P. The biology of peroxisome proliferator-activated receptors: relationship with lipid metabolism and insulin sensitivity. Diabetes. 2004 Feb;53 Suppl 1S43-S50. 21. Delarue J, LeFoll C, Corporeau C, Lucas D. N-3 long chain polyunsaturated fatty acids: a nutritional tool to prevent insulin resistance associated to type 2 diabetes and obesity? Reprod Nutr Dev. 2004 May;44(3):289-99. 22. Li H, Ruan XZ, Powis SH, et al. EPA and DHA reduce LPS-induced inflammation responses in HK-2 cells: evidence for a PPAR-gamma-dependent mechanism. Kidney Int. 2005 Mar;67(3):867-74. 23. Madsen T, Skou HA, Hansen VE, et al. C-reactive protein, dietary n-3 fatty acids, and the extent of coronary artery disease. Am J Cardiol. 2001 Nov 15;88(10):1139-42. 24. Mori TA, Puddey IB, Burke V, et al. Effect of omega 3 fatty acids on oxidative stress in humans: GC-MS measurement of urinary F2-isoprostane excretion. Redox Rep. 2000;5(1):45-6. 25. Cho KJ, Moon HE, Moini H, et al. Alpha-lipoic acid inhibits adipocyte differentiation by regulating pro-adipogenic transcription factors via mitogen-activated protein kinase pathways. J Biol Chem. 2003 Sep 12;278(37):34823-33. 26. Jacob S, Ruus P, Hermann R, et al. Oral administration of RAC-alpha-lipoic acid modulates insulin sensitivity in patients with type-2 diabetes mellitus: a placebo-controlled pilot trial. Free Radic Biol Med. 1999 Aug;27(3-4):309-14. 27. Packer L, Kraemer K, Rimbach G. Molecular aspects of lipoic acid in the prevention of diabetes complications. Nutrition. 2001 Oct;17(10):888-95. 28. Sola S, Mir MQ, Cheema FA, et al. Irbesartan and lipoic acid improve endothelial function and reduce markers of inflammation in the metabolic syndrome: results of the Irbesartan and Lipoic Acid in Endothelial Dysfunction (ISLAND) study. Circulation. 2005 Jan 25;111(3):343-8. 29. Dentin R, Pegorier JP, Benhamed F, et al. Hepatic glucokinase is required for the synergistic action of ChREBP and SREBP-1c on glycolytic and lipogenic gene expression. J Biol Chem. 2004 May 7;279(19):20314-26. 30. Chen YD, Coulston AM, Zhou MY, Hollenbeck CB, Reaven GM. Why do low-fat high-carbohydrate diets accentuate postprandial lipemia in patients with NIDDM? Diabetes Care. 1995 Jan;18(1):10-6. 31. Esposito K, Marfella R, Ciotola M, et al. Effect of a mediterranean-style diet on endothelial dysfunction and markers of vascular inflammation in the metabolic syndrome: a randomized trial. JAMA. 2004 Sep 22;292(12):1440-6. 32. Singh RB, Dubnov G, Niaz MA, et al. Effect of an Indo-Mediterranean diet on progression of coronary artery disease in high risk patients (Indo-Mediterranean Diet Heart Study): a randomised single-blind trial. Lancet. 2002 Nov 9;360(9344):1455-61. 33. Wien MA, Sabate JM, Ikle DN, Cole SE, Kandeel FR. Almonds vs complex carbohydrates in a weight reduction program. Int J Obes Relat Metab Disord. 2003 Nov;27(11):1365-72. 34. Savage DB, Petersen KF, Shulman GI. Mechanisms of insulin resistance in humans and possible links with inflammation. Hypertension. 2005 May;45(5):828-33. 35. Perseghin G, Petersen K, Shulman GI. Cellular mechanism of insulin resistance: potential links with inflammation. Int J Obes Relat Metab Disord. 2003 Dec;27 Suppl 3S6-11. 36. Comalada M, Camuesco D, Sierra S, et al. In vivo quercitrin anti-inflammatory effect involves release of quercetin, which inhibits inflammation through down-regulation of the NF-kappaB pathway. Eur J Immunol. 2005 Feb;35(2):584-92. 37. Donnelly LE, Newton R, Kennedy GE, et al. Anti-inflammatory effects of resveratrol in lung epithelial cells: molecular mechanisms. Am J Physiol Lung Cell Mol Physiol. 2004 Oct;287(4):L774-83. 38. Carluccio MA, Siculella L, Ancora MA, et al. Olive oil and red wine antioxidant polyphenols inhibit endothelial activation: antiatherogenic properties of Mediterranean diet phytochemicals. Arterioscler Thromb Vasc Biol. 2003 Apr 1;23(4):622-9. 39. Martinez-Dominguez E, de la PR, Ruiz-Gutierrez V. Protective effects upon experimental inflammation models of a polyphenol-supplemented virgin olive oil diet. Inflamm Res. 2001 Feb;50(2):102-6. 40. Wiseman SA, Mathot JN, de Fouw NJ, Tijburg LB. Dietary non-tocopherol antioxidants present in extra virgin olive oil increase the resistance of low density lipoproteins to oxidation in rabbits. Atherosclerosis. 1996 Feb;120(1-2):15-23. 41. Sies H, Stahl W, Sevanian A. Nutritional, dietary and postprandial oxidative stress. J Nutr. 2005 May;135(5):969-72. 42. Azzi A, Gysin R, Kempna P, et al. Regulation of gene expression by alpha-tocopherol. Biol Chem. 2004 Jul;385(7):585-91. 43. Fattoretti P, Bertoni-Freddari C, Casoli T, et al. Decreased expression of glucose transport protein (Glut3) in aging and vitamin E deficiency. Ann NY Acad Sci. 2002 Nov;973:293-6. 44. Ford ES, Mokdad AH, Giles WH, Brown DW. The metabolic syndrome and antioxidant concentrations: findings from the Third National Health and Nutrition Examination Survey. Diabetes. 2003 Sep;52(9):2346-52. 45. Felipe F, Bonet ML, Ribot J, Palou A. Modulation of resistin expression by retinoic acid and vitamin A status. Diabetes. 2004 Apr;53(4):882-9. 46. Matthews KA, Rhoten WB, Driscoll HK, Chertow BS. Vitamin A deficiency impairs fetal islet development and causes subsequent glucose intolerance in adult rats. J Nutr. 2004 Aug;134(8):1958-63. 47. Khan A, Safdar M, Ali Khan MM, Khattak KN, Anderson RA. Cinnamon improves glucose and lipids of people with type 2 diabetes. Diabetes Care. 2003 Dec;26(12):3215-8. 48. Mancini-Filho J, Van Koiij A, Mancini DA, Cozzolino FF, Torres RP. Antioxidant activity of cinnamon (Cinnamomum Zeylanicum, Breyne) extracts. Boll Chim Farm. 1998 Dec;137(11):443-7. 49. Anderson RA, Broadhurst CL, Polansky MM, et al. Isolation and characterization of polyphenol type-A polymers from cinnamon with insulin-like biological activity. J Agric Food Chem. 2004 Jan 14;52(1):65-70. 50. Imparl-Radosevich J, Deas S, Polansky MM, et al. Regulation of PTP-1 and insulin receptor kinase by fractions from cinnamon: implications for cinnamon regulation of insulin signalling. Horm Res. 1998 Sep;50(3):177-82. 51. Jarvill-Taylor KJ, Anderson RA, Graves DJ. A hydroxychalcone derived from cinnamon functions as a mimetic for insulin in 3T3-L1 adipocytes. J Am Coll Nutr. 2001 Aug;20(4):327-36. 52. Selivankina SY, Zubo YO, Zubkova NK, et al. Cytokinin control of rRNA gene transcription in senescing barley leaves. Dokl Biochem Biophys. 2004 Jul;397:258-60. 53. Galuszka P, Frebortova J, Werner T, et al. Cytokinin oxidase/dehydrogenase genes in barley and wheat: cloning and heterologous expression. Eur J Biochem. 2004 Oct;271(20):3990-4002. 54. Hong H, Jai MW. Effects of malted barley extract and banaba extract on blood glucose levels in genetically diabetic mice. J Med Food. 2004;7(4):487-90. 55. Available at: http://www.futureceuticals.com. Accessed September 6, 2005. 56. Tuomilehto J, Hu G, Bidel S, Lindstrom J, Jousilahti P. Coffee consumption and risk of type 2 diabetes mellitus among middle-aged Finnish men and women. JAMA. 2004 Mar 10;291(10):1213-9. 57. Salazar-Martinez E, Willett WC, Ascherio A, et al. Coffee consumption and risk for type 2 diabetes mellitus. Ann Intern Med. 2004 Jan 6;140(1):1-8. 58. Yen WJ, Wang BS, Chang LW, Duh PD. Antioxidant properties of roasted coffee residues. J Agric Food Chem. 2005 Apr 6;53(7):2658-63. 59. Rodriguez de Sotillo DV, Hadley M. Chlorogenic acid modifies plasma and liver concentrations of: cholesterol, triacylglycerol, and minerals in (fa/fa) Zucker rats. J Nutr Biochem. 2002 Dec;13(12):717-26. 60. Johnston KL, Clifford MN, Morgan LM. Coffee acutely modifies gastrointestinal hormone secretion and glucose tolerance in humans: glycemic effects of chlorogenic acid and caffeine. Am J Clin Nutr. 2003 Oct;78(4):728-33. 61. McCarty MF. A chlorogenic acid-induced increase in GLP-1 production may mediate the impact of heavy coffee consumption on diabetes risk. Med Hypotheses. 2005;64(4):848-53. 62. Ochiai R, Jokura H, Suzuki A, et al. Green coffee bean extract improves human vasoreactivity. Hypertens Res. 2004 Oct;27(10):713-7. |



Wellness
Specialists
1-800-226-2370 - This service is FREE
7:30 AM - 12 AM (ET) Mon-Fri | 9 AM - 12 AM (ET) Sat-Sun