Life Extension Magazine®

Why Does Your Body Need Citrus Fruits
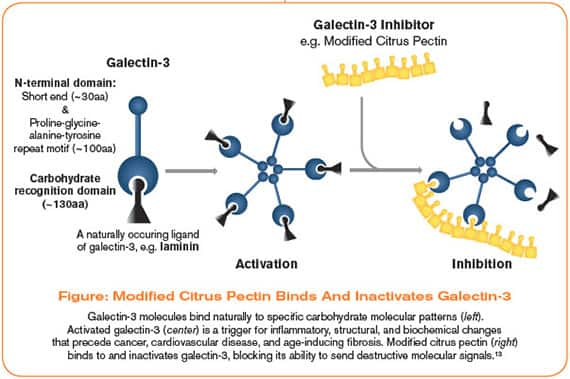
A little-known molecule produced in our bodies called galectin-3 has been found to play a causative role in heart failure, cancer, and kidney disease. As drug companies search for a solution, scientists have discovered that modified citrus pectin, derived from the pith of citrus fruits, can inhibit galectin-3, blocking its ability to send destructive molecular signals.
Scientifically reviewed by: Dr. Shaylind Benson, ND, in October 2024. Written by: Steven De Berg.

There are molecules in our bodies that are beneficial in youth, but turn against us as we age.
One of these compounds is galectin-3. Excess levels are associated with heart failure, kidney disease, and cancer.1-4
Doctors have long recognized galectin-3 as a biomarker for degenerative disease.5-10 New research is revealing that galectin-3 is far more than an indicator of disease—it is purported to have a causative role in these conditions.11-14
Drug companies have had little success in their attempts to fight the destructive actions of galectin-3. Fortunately, scientists have discovered a citrus extract that has potent galectin-3 inhibitory properties.
Found in the pith of citrus fruit peels, modified citrus pectin (MCP) has been shown to inactivate galectin-3, blocking its ability to send destructive molecular signals throughout the body.4
Because of its ability to block galectin-3, modified citrus pectin is emerging as a key natural compound in the battle against heart failure, cancer, and kidney disease.4,15,16
Keeping Galectin-3 In Balance

Galectin-3, like many biologically active molecules, has a distinct and important role in maintaining good health, but galectin-3 becomes a problem when it occurs in excessive amounts. In normal health, for example, galectin-3 provides an important defense against bacteria following acute tissue damage by helping induce local inflammation and an immune response that destroys the invading organisms.17 And with chronic or repetitive tissue injury, galectin-3 participates in the transition to chronic inflammation, helping to “wall off” the injured or infected area.17
The problem with this action is that it produces tissue scarring that in the long run impairs the function of the very organ or tissue it was trying to protect.17 In addition, galectin-3 is involved in an important process called “tissue remodeling,” in which cells in various tissues become altered in response to damaging stimuli. Remodeling is an important part of heart failure and stroke,18-21 and although we might not typically think of it this way, cancer can be viewed as an aggressive and malignant form of remodeling.22
These multiple actions mean that galectin-3 has a wide variety of biological functions and plays a key role in physiological and pathological processes involved in cancer, cardiovascular disease, and the pro-aging process of fibrosis.17,23-25 All of these disorders share a common factor of increased inflammation and altered immune function, so galectin-3 may be considered a pro-inflammatory, immune-altering molecule.26,27 In addition, levels of galectin-3 rise significantly with advancing age, making it a prime target for anti-aging therapies.25,28
The Role of Galectin-3 In Cancer Development
For many cancers, rates rise dramatically with advancing age.29,30 There is now plenty of evidence that some of the hallmarks of developing cancers, such as inflammation, immune function alterations, and local and metastatic spread, are related to galectin-3 actions.
Because increased concentrations of circulating galectin-3 are so prevalent in growing cancers, it has been suggested as a biomarker for specific cancers, such as prostate, ovarian, and breast cancer.31-34 But increasing evidence suggests that galectin-3 does more than signal the presence of cancer—it actively participates in cancer growth by promoting cancer cell adhesion to blood vessel linings, a major step in the progression and spread of cancer.1,2
In addition, the excess galectin-3 produced by cancer cells helps the malignant cells communicate, stick together, rapidly multiply, grow necessary new blood vessels, and evade the normal programmed cell death called apoptosis.18,26,35,36
Recently, it has been demonstrated that galectin-3 enables cell migration and invasion of melanoma-induced metastasis to the lungs by inducing secretion of matrix metalloproteinases (MMP-1 and MMP-9) required to breach vascular basement membranes to the lungs.37,38 It was shown that galectin-3 is expressed on all the major compartments of the lungs and participates in not just promoting adhesion but also in spreading the cancer to lungs. Additional laboratory studies suggest that overexpression of galectin-3 in lung tissue increases cancer cell adhesion, motility, and organ colonization.39,40
Galectin-3’s Role In Heart Failure
Studies show that galectin-3 plays a major role in heart failure.
Galectin-3 is a biomarker for fibrosis and inflammation in heart failure.3 People hospitalized for heart failure with galectin-3 levels exceeding 17.8 ng/mL have 2.8 times the odds of being re-admitted at 30 days, and more than three times the odds of being readmitted by 90 days, compared with those having galectin-3 levels below this value.41
In one study, patients whose galectin-3 levels doubled over 18 months were nearly two times as likely to be hospitalized for heart failure and die of any of a number of causes.42 That study also showed a close association between galectin-3 levels and inflammatory markers such as interleukin-6 (IL-6) and C-reactive protein (CRP). Other studies have demonstrated a close correlation between galectin-3 levels and blood pressure, serum lipids, body mass index, kidney function, and other biological markers of disease.3,18
However, as is the case with cancer, galectin-3 is now recognized as being more than simply a biomarker of impending heart failure—it’s now believed to play a major causative role in the development of heart disease.
Failing heart tissue has been found to produce increased quantities of galectin-3 before clinically evident heart failure.25,27 In advanced cases of heart failure, galectin-3 levels have been correlated with many of the physical changes that occur as the heart progressively becomes less able to pump blood, such as thickening of the heart wall.27
Animal studies show that increased expression of galectin-3 produces heart muscle dysfunction, with a massive overproduction of the tough, unyielding form of connective tissue called type I collagen that impairs heart muscle elasticity.15,43 This loss of elasticity and the decreased pumping action that follows are hallmarks of heart failure.
Similarly, animal studies have shown that galectin-3 is found in increased amounts in brain tissue following an ischemic (lack of blood flow) stroke, playing a role in post-stroke tissue remodeling and abnormal new blood vessel formation. 18
Indeed, galectin-3 is now known to play a causative role in the remodeling of heart and blood vessel tissue that occurs during heart failure.18,44 This remodeling is linked closely to disease progression and poor prognosis in heart failure. Galectin-3 stimulates the migration of inflammatory cells, proliferation of fibroblasts (the tough, scar-like cells that replace normal contractile heart muscle), and ultimately the development of fibrosis, the stiffening process that impairs heart muscle function.45
What You Need To Know
 |
Modified Citrus Pectin
- A common feature of cancer, heart failure, and kidney disease is the pathological replication and reorganization of healthy cells, manifested as changes in tissue structure.
- This harmful “remodeling” of tissue has recently been found to be under the control of a unique molecule called galectin-3.
- Higher galectin-3 levels have now been associated with increased risk and severity of cancer, greater severity and more rapid progression of heart failure, and with both acute and chronic kidney disease.
- Modified citrus pectin (MCP) binds to and inactivates galectin-3, preventing it from delivering its destructive signals to cells and tissues.
- Studies now demonstrate the effectiveness of MCP in laboratory and preclinical settings, where it counteracts galectin-3 actions, slows tissue remodeling, and reduces consequences such as cancer development and metastasis, heart failure, and kidney disease.
- Big Pharma is hot on the trail of drugs that block galectin-3, but MCP is available now in convenient dosing forms, with no recognized side effects.
Nature’s Answer To High Galectin-3 Levels
As the connection between galectin-3 and these major diseases becomes better known, Big Pharma has been hotly pursuing drugs that combat disease by blocking galectin-3. So far it has come up short, but as usual, nature already has a solution. Found in the pith of citrus fruit peels, a compound called modified citrus pectin (MCP) has been shown to inactivate galectin-3.
Most people have heard of pectin as a gelling agent used in fruit preserves and jellies. In fact, many of the chemical properties that make pectin appealing in cooking are similar to those that attract the attention of scientists. Modified citrus pectin is a form of pectin that is especially rich in sugar molecules known as galactosides.16,46
Galectin-3 molecules specifically interact with the galactosides found in MCP (modified citrus pectin).4 This means that, in the presence of this special type of pectin, galectin-3 molecules will bind to the galactoside bonds on the MCP molecules, rather than on those signaling carbohydrate molecules on cell surfaces.47
In this way, pectin acts as a “competitive inhibitor,” binding up and inactivating galectin-3, and preventing it from performing the actions that can damage your health. (See Figure below).4,13,16
Laboratory and clinical evidence has demonstrated that modified citrus pectin’s ability to inhibit galectin-3 helps prevent the formation and spread of cancer, helps mitigate cardiovascular disease, and generally contributes to fighting the pro-aging effects of fibrosis in your body.
Modified Citrus Pectin Diminishes Cancer Risk
Leading cancer researchers have described MCP as “one bullet, multiple targets” because of its multiple, complementary actions in fighting cancer.48
Modified citrus pectin helps prevent the spread of cancer.
Modified citrus pectin has demonstrated tremendous anticancer potential, particularly in fighting deadly metastatic spread. This is important because once a cancer metastasizes (spreads to a distant part of the body), it becomes vastly more dangerous and difficult to treat.16
Early preclinical studies showed that injecting mice with MCP could significantly decrease the likelihood of melanoma (a deadly skin cancer) spreading to the lungs.46,48,49 The mechanism was later shown to be related to the binding of MCP to galectin-3, which inhibited malignant cells’ ability to grow and stick to normal tissue and blood vessels; this “stickiness” is now recognized as an essential part of the formation of metastases.50 Recent research has shown that metastasis of melanoma to the lungs could be hindered by feeding mice MCP in their drinking water, inhibiting galectin-3 found on the lung tissue. The authors concluded that MCP competes with the carbohydrate found on melanoma cells (polyLacNAc) for binding to galectin-3 on the lungs, thereby impacting metastasis.38
Similar antimetastatic actions were also shown for orally administered MCP in animal models of both prostate cancer (which becomes very difficult to treat after it metastasizes) and breast cancer.50-54
Further preclinical studies showed that by binding to galectin-3, MCP prevented colon cancer from spreading to the liver.55 The same model showed that MCP reduces colon cancer size by up to 70% compared with controls.56
What Is Galectin-3?
Galectin-3 is a member of a family of biological compounds responsible for recognizing and responding to specific patterns found on carbohydrate molecules.23,24,71 Many vital cells in the body come ready “labeled” with those specific carbohydrate patterns, so galectin-3 functions as a means for cells to recognize and interact with one another and with the “matrix” material that supports cells in their correct positions.24,35 Galectin-3 has a particular molecular region that has an affinity for the sugar galactose and its bonds to adjacent molecules, which are called galactoside bonds.26
Galectin-3 levels vary by cell type and function; the molecule is secreted from cells in a unique and not well-understood fashion.23,26 Galectin-3 functions as an autocrine/paracrine hormone when present outside the cells.72 One important feature of galectin-3 is its ability to form small clusters of galectin-3 molecules bound together in pentamers. This allows the molecule to participate in multiple cellular and extracellular processes involving tissue growth, structure, and immune modulation.17,72,73
Modified citrus pectin activates cancer-killing cells.
Studies on human blood samples reveal that MCP also activates cancer-killing immune system cells (natural killer, or NK, cells), boosting their ability to patrol for and eliminate cancers as soon as they form; this is a boost to one of the body’s most important anticancer mechanisms.57
Modified citrus pectin induces apoptosis.
In addition to helping to prevent metastases, research on prostate cancer cells shows that MCP works earlier in the production of cancer by restoring malignant cells’ ability to die off normally through programmed cell death (apoptosis).58 Importantly, these anticancer actions are effective for both hormone-dependent and hormone-independent cancers; the latter are notoriously challenging to treat.58,59
In humans with prostate cancer, MCP has been shown to prolong the time it takes for doubling of prostate-specific antigen (PSA) levels; the more slowly PSA doubles, the more slowly the tumor is growing.60
Modified citrus pectin helps boost the effectiveness of cancer treatments.
In laboratory studies of human multiple myeloma (a cancer that affects certain types of white blood cells) and angiosarcoma (a deadly cancer of blood vessels), MCP helps make malignant cells more responsive to chemotherapy while increasing cell death by apoptosis.61-63
Similar chemo-sensitizing actions of MCP have now been shown for other cancers as well, including prostate and ovarian cancers.33,64,65 In a laboratory study, MCP was also recently shown to synergize with two natural biological mixtures to fight breast and prostate cancer metastases.66
How MCP Battles Galectin-3 In Heart Disease

Heart failure remains one of the most challenging problems in clinical medicine today.
It is now known that 30 to 50% of patients with increased galectin-3 levels have progressive or remodeling form of heart failure.13
Once thought to be simply a marker of severe, progressive heart failure, galectin-3 is now thought to play a major role in triggering the remodeling and poor function seen in the failing heart.12,13,27,67-69
The good news is that binding or inhibiting galectin-3 can prevent and may even reverse heart failure, even heart failure with extensive fibrosis.67
Laboratory studies show that MCP directly counteracts galectin-3-induced changes in heart muscle structure and function.
In one study, galectin-3 was shown to increase production of type I collagen, a major component in arterial stiffness and heart failure; however, inhibiting galectin-3 with modified citrus pectin blocked synthesis of type I collagen.15 In this study, when hypertensive rats were treated with the hormone aldosterone (which increases blood pressure and worsens heart failure), the control animals demonstrated thickening of blood vessel walls, inflammation, fibrosis, and increased expression of galectin-3; however, treatment with modified citrus pectin reversed all of these effects.15
One of the underlying risk factors for developing heart attacks, stroke, and heart failure is atherosclerosis, the “hardening” or thickening of the arteries.70 Atherosclerotic plaques, the bulging masses of cholesterol and inflammatory cells that threaten to block arteries, are laden with high levels of galectin-3.70 When mice that developed atherosclerosis as a result of a high-cholesterol “Western” diet were given oral doses of modified citrus pectin, the volume of their plaques was diminished, the result of galectin-3 inhibition.70
Recent studies have highlighted the importance of measuring galectin-3 levels in people at high risk for cardiovascular disease, particularly heart failure and cardiovascular fibrosis.13,18,42,45 As the evidence grows that galectin-3 is an active participant, not an innocent bystander or simple disease marker, the importance of inhibiting its damaging effects on the heart will be amplified. Modified citrus pectin may offer a simple, natural, safe route to minimizing the impact of galectin-3 on your heart.
MCP Fights Kidney Disease And Fibrosis
Kidney disease and acute kidney injuries involve increased levels and activity of galectin-3. In an animal model of acute kidney injury, the damaged kidneys increased in size, and galectin-3 expression was widespread in several types of tubules inside the kidney.4
Research in mice show that when modified citrus pectin is administered before kidney damage is induced experimentally, it significantly reduces kidney enlargement and cuts down on kidney cell proliferation, an early step in fibrosis.4 Once the damaging stimulus was removed, MCP-treated mice showed decreased fibrosis, fewer inflammatory cells and cytokines in kidney tissue, and increased programmed cell death (apoptosis)—all in association with reduced levels of galectin-3.4
This study demonstrates how modified citrus pectin not only opposes the immediate damaging effects of galectin-3, but also reduces its levels, making for a vastly healthier kidney environment.
Summary
Galectin-3 is a signaling molecule that is involved in tissue growth and repair. This is beneficial in youth, but galectin-3 levels rise with age, and by midlife, galectin-3 may represent more of a threat than an asset. Higher galectin-3 levels serve as markers for elevated cancer risk, cardiovascular disease risk and severity, and kidney disease.
Galectin-3, however, is not simply a marker of disease. Scientists have now shown that galectin-3 is an active player, triggering the harmful changes that characterize each of these conditions. Studies show that inhibiting galectin-3 can markedly reduce, and in some cases reverse, dangerous tissue changes induced by the molecule.
Modified citrus pectin (MCP) is a natural product that inhibits galectin-3, shutting down its ability to communicate with target cells. In the presence of MCP, cancer cells lose their heightened survival and reproductive abilities, as well as their capacity to spread (metastasis); heart tissue undergoes less of the dangerous remodeling that is typical of heart failure following a heart attack or sustained stresses from high blood pressure; and kidney cells become resistant to formation of fibrosis that impairs kidney function.
The future likely holds much more promise for modified citrus pectin as we gain knowledge about galectin-3 and its ubiquitous role in disease. For now, it makes sense for certain individuals to add this supplement to control aberrant galectin-3.
A typical high-dose is to take five grams of MCP powder three times a day for several months, and then reduce to five grams once a day. Some people will only take MCP during specific time periods, and then discontinue.
MCP should be taken away from meals, i.e. on an empty or almost empty stomach.
MCP is not yet recommended as a daily supplement for everyone to take. Certain individuals will take the full 15 gram/day dose for around one year, while others may try a moderate dose course (5 grams/day) for a period of 2-4 months.
If you have any questions on the scientific content of this article, please call a Life Extension® Health Advisor at 1-866-864-3027.
References
- Yu LG, Andrews N, Zhao Q, et al. Galectin-3 interaction with Thomsen-Friedenreich disaccharide on cancer-associated MUC1 causes increased cancer cell endothelial adhesion. J Biol Chem. 2007 Jan 5;282(1):773-81.
- Zhao Q, Guo X, Nash GB, et al. Circulating galectin-3 promotes metastasis by modifying MUC1 localization on cancer cell surface. Cancer Res. 2009 Sep 1;69(17):6799-806.
- Lok DJ, Van Der Meer P, de la Porte PW, et al. Prognostic value of galectin-3, a novel marker of fibrosis, in patients with chronic heart failure: data from the DEAL-HF study. Clin Res Cardiol. 2010 May;99(5):323-8.
- Kolatsi-Joannou M, Price KL, Winyard PJ, Long DA. Modified citrus pectin reduces galectin-3 expression and disease severity in experimental acute kidney injury. PLoS One. 2011;6(4):e18683.
- Falcone C, Lucibello S, Mazzucchelli I, Bozzini S, D’Angelo A, Schirinzi S, et al. Galectin-3 plasma levels and coronary artery disease: a new possible biomarker of acute coronary syndrome. Int J Immunopathol Pharmacol. 2011 Oct-Dec;24(4):905-13.
- Sherwi N, Merali S, Wong K. Personalizing biomarker strategies in heart failure with galectin-3. Future Cardiol. 2012 Nov;8(6):885-94.
- O’Seaghdha CM, Hwang SJ, Ho JE, Vasan RS, Levy D, Fox CS. Elevated galectin-3 precedes the development of CKD. J Am Soc Nephrol. 2013 Sep;24(9):1470-7.
- Balan V, Nangia-Makker P, Raz A. Galectins as cancer biomarkers. Cancers (Basel). 2010 Apr 20;2(2):592-610.
- Chiu CG, Strugnell SS, Griffith OL, Jones SJ, Gown AM, Walker B, et al. Diagnostic utility of galectin-3 in thyroid cancer. Am J Pathol. 2010 May;176(5):2067-81.
- Gopal DM, Kommineni M, Ayalon N, Koelbl C, Ayalon R, Biolo A, et al. Relationship of plasma galectin-3 to renal function in patients with heart failure: effects of clinical status, pathophysiology of heart failure, and presence or absence of heart failure. J Am Heart Assoc. 2012 Oct;1(5):e000760. Erratum in J Am Heart Assoc. 2013 Jun;2(3):e000194.
- de Boer RA, Edelmann F, Cohen-Solal A, Mamas MA, Maisel A, Pieske B. Galectin-3 in heart failure with preserved ejection fraction. Eur J Heart Fail. 2013 Oct;15(10):1095-101.
- Cecchinelli B, Lavra L, Rinaldo C, Iacovelli S, Gurtner A, Gasbarri A, et al. Repression of the antiapoptotic molecule galectin-3 by homeodomain-interacting protein kinase 2-activated p53 is required for p53-induced apoptosis. Mol Cell Biol. 2006 Jun;26(12):4746-57.
- de Boer RA, van der Velde AR, Mueller C, van Veldhuisen DJ, Anker SD, Peacock WF, et al. Galectin-3: a modifiable risk factor in heart failure. Cardiovasc Drugs Ther. 2014 Jun;28(3):237-46.
- Ho JE, Liu C, Lyass A, Courchesne P, Pencina MJ, Vasan RS, et al. Galectin-3, a marker of cardiac fibrosis, predicts incident heart failure in the community. J Am Coll Cardiol. 2012 Oct 2;60(14):1249-56.
- Calvier L, Miana M, Reboul P, et al. Galectin-3 mediates aldosterone-induced vascular fibrosis. Arterioscler Thromb Vasc Biol. 2013 Jan;33(1):67-75.
- Modified citrus pectin-monograph. Altern Med Rev. 2000 Dec;5(6):573-5.
- Henderson NC, Sethi T. The regulation of inflammation by galectin-3. Immunol Rev. 2009 Jul;230(1):160-71.
- Yan YP, Lang BT, Vemuganti R, Dempsey RJ. Galectin-3 mediates post-ischemic tissue remodeling. Brain Res. 2009 Sep 8;1288:116-24.
- Konstam MA, Kramer DG, Patel AR, Maron MS, Udelson JE. Left ventricular remodeling in heart failure: current concepts in clinical significance and assessment. JACC Cardiovasc Imaging. 2011 Jan;4(1):98-108.
- Weber KT. Extracellular matrix remodeling in heart failure: a role for de novo angiotensin II generation. Circulation. 1997 Dec 2;96(11):4065-82.
- Ellison JA, Barone FC, Feuerstein GZ. Matrix remodeling after stroke. De novo expression of matrix proteins and integrin receptors. Ann N Y Acad Sci. 1999;890:204-22.
- Markert EK, Levine AJ, Vazquez A. Proliferation and tissue remodeling in cancer: the hallmarks revisited. Cell Death Dis. 2012 Oct 4;3:e397.
- Krzeslak A, Lipinska A. Galectin-3 as a multifunctional protein. Cell Mol Biol Lett. 2004;9(2):305-28.
- Dumic J, Dabelic S, Flogel M. Galectin-3: an open-ended story. Biochim Biophys Acta. 2006 Apr;1760(4):616-35.
- de Boer RA, van Veldhuisen DJ, Gansevoort RT, et al. The fibrosis marker galectin-3 and outcome in the general population. J Intern Med. 2012 Jul;272(1):55-64.
- Chen HY, Liu FT, Yang RY. Roles of galectin-3 in immune responses. Arch Immunol Ther Exp (Warsz). 2005 Nov-Dec;53(6):497-504.
- Sharma UC, Pokharel S, van Brakel TJ, et al. Galectin-3 marks activated macrophages in failure-prone hypertrophied hearts and contributes to cardiac dysfunction. Circulation. 2004 Nov 9;110(19):3121-8.
- Shah RV, Chen-Tournoux AA, Picard MH, van Kimmenade RR, Januzzi JL. Galectin-3, cardiac structure and function, and long-term mortality in patients with acutely decompensated heart failure. Eur J Heart Fail. 2010 Aug;12(8):826-32.
- Howlader N, Noone AM, Krapcho M, Garshell J, Miller D, Altekruse SF, et al.(eds). SEER Cancer Statistics Review, 1975-2011, National Cancer Institute. Bethesda, MD.
- Thakkar JP, McCarthy BJ, Villano JL. Age-specific cancer incidence rates increase through the oldest age groups. Am J Med Sci. 2014 Jul;348(1):65-70.
- Balan V, Wang Y, Nangia-Makker P, et al. Galectin-3: a possible complementary marker to the PSA blood test. Oncotarget. 2013 Apr;4(4):542-9.
- Eliaz I. The role of galectin-3 as a marker of cancer and inflammation in a stage IV ovarian cancer patient with underlying pro-inflammatory comorbidities. Case Rep Oncol. 2013 May;6(2):343-9.
- Hossein G, Keshavarz M, Ahmadi S, Naderi N. Synergistic effects of PectaSol-C modified citrus pectin an inhibitor of Galectin-3 and paclitaxel on apoptosis of human SKOV-3 ovarian cancer cells. Asian Pac J Cancer Prev. 2013;14(12):7561-8.
- Yu LG. Circulating galectin-3 in the bloodstream: An emerging promoter of cancer metastasis. World J Gastrointest Oncol. 2010 Apr 15;2(4):177-80.
- Califice S, Castronovo V, Van Den Brule F. Galectin-3 and cancer (review). Int J Oncol. 2004 Oct;25(4):983-92.
- Newlaczyl AU, Yu LG. Galectin-3—a jack-of-all-trades in cancer. Cancer Lett. 2011 Dec 27;313(2):123-8.
- Wang YG, Kim SJ, Baek JH, Lee HW, Jeong SY, Chun KH. Galectin-3 increases the motility of mouse melanoma cells by regulating matrix metalloproteinase-1 expression. Exper Mol Med. 2012;44(6):387-93.
- Dange MC, Srinivasan N, More SK, et al. Galectin-3 expressed on different lung compartments promotes organ specific metastasis by facilitating arrest, extravasation and organ colonization via high affinity ligands on melanoma cells. Clin Exp Metastasis. 2014 Jun 21. [Epub ahead of print]
- O’Driscoll L, Linehan R, Liang YH, Joyce H, Oglesby I, Clynes M. Galectin-3 expression alters adhesion, motility and invasion in a lung cell line (DKLP), in vitro. Anticancer Res. 2002 Nov-Dec;22(6A):3117-25.
- Buttery R, Monaghan H, Salter DM, Sethi T. Galectin-3: differential expression between small-cell and non-small-cell lung cancer. Histopathology. 2004 Apr;44(4):339-44.
- Meijers WC, Januzzi JL, deFilippi C, et al. Elevated plasma galectin-3 is associated with near-term rehospitalization in heart failure: A pooled analysis of 3 clinical trials. Am Heart J. 2014 Jun;167(6):853-60.e4.
- de Boer RA, Lok DJ, Jaarsma T, et al. Predictive value of plasma galectin-3 levels in heart failure with reduced and preserved ejection fraction. Ann Med. 2011 Feb;43(1):60-8.
- Wehrens XH, Marks AR. Molecular determinants of altered contractility in heart failure. Ann Med. 2004;36 Suppl 1:70-80.
- Xue Y, Maisel A, Peacock WF. Using galectin-3 to reduce heart failure rehospitalization. Future Cardiol. 2014 Mar;10(2):221-7.
- de Boer RA, Voors AA, Muntendam P, van Gilst WH, van Veldhuisen DJ. Galectin-3: a novel mediator of heart failure development and progression. Eur J Heart Fail. 2009 Sep;11(9):811-7.
- Inohara H, Raz A. Effects of natural complex carbohydrate (citrus pectin) on murine melanoma cell properties related to galectin-3 functions. Glycoconj J. 1994 Dec;11(6):527-32.
- Gao X, Zhi Y, Sun L, et al. The inhibitory effects of a rhamnogalacturonan I (RG-I) domain from ginseng pectin on galectin-3 and its structure-activity relationship. J Biol Chem. 2013 Nov 22;288(47):33953-65.
- Glinsky VV, Raz A. Modified citrus pectin anti-metastatic properties: one bullet, multiple targets. Carbohydr Res. 2009 Sep 28;344(14):1788-91.
- Platt D, Raz A. Modulation of the lung colonization of B16-F1 melanoma cells by citrus pectin. J Natl Cancer Inst. 1992 Mar 18;84(6):438-42.
- Glinskii OV, Huxley VH, Glinsky GV, Pienta KJ, Raz A, Glinsky VV. Mechanical entrapment is insufficient and intercellular adhesion is essential for metastatic cell arrest in distant organs. Neoplasia. 2005 May;7(5):522-7.
- Pienta KJ, Naik H, Akhtar A, et al. Inhibition of spontaneous metastasis in a rat prostate cancer model by oral administration of modified citrus pectin. J Natl Cancer Inst. 1995 Mar 1;87(5):348-53.
- Glinskii OV, Sud S, Mossine VV, Mawhinney TP, Anthony DC, Glinsky GV, et al. Inhibition of prostate cancer bone metastasis by synthetic TF antigen mimic/galectin-3 inhibitor lactulose-L-leucine. Neoplasia. 2012 Jan;14(1):65-73.
- Ye L, Kynaston HG, Jiang WG. Bone metastasis in prostate cancer: molecular and cellular mechanisms (review). Int J Mol Med. 2007 Jul;20(1):103-11.
- Mohammad KS, Fournier PG, Guise TA, Chirgwin JM. Agents targeting prostate cancer bone metastasis. Anticancer Agents Med Chem. 2009 Dec;9(10):1079-88.
- Liu HY, Huang ZL, Yang GH, Lu WQ, Yu NR. Inhibitory effect of modified citrus pectin on liver metastases in a mouse colon cancer model. World J Gastroenterol. 2008 Dec 28;14(48):7386-91.
- Hayashi A, Gillen AC, Lott JR. Effects of daily oral administration of quercetin chalcone and modified citrus pectin on implanted colon-25 tumor growth in Balb-c mice. Altern Med Rev. 2000 Dec;5(6):546-52.
- Ramachandran C, Wilk BJ, Hotchkiss A, Chau H, Eliaz I, Melnick SJ. Activation of human T-helper/inducer cell, T-cytotoxic cell, B-cell, and natural killer (NK)-cells and induction of natural killer cell activity against K562 chronic myeloid leukemia cells with modified citrus pectin. BMC Complement Altern Med. 2011;11:59.
- Yan J, Katz A. PectaSol-C modified citrus pectin induces apoptosis and inhibition of proliferation in human and mouse androgen-dependent and- independent prostate cancer cells. Integr Cancer Ther. 2010 Jun;9(2):197-203.
- Oberstein PE, Olive KP. Pancreatic cancer: why is it so hard to treat? Therap Adv Gastroenterol. Jul 2013;6(4):321-37.
- Guess BW, Scholz MC, Strum SB, Lam RY, Johnson HJ, Jennrich RI. Modified citrus pectin (MCP) increases the prostate-specific antigen doubling time in men with prostate cancer: a phase II pilot study. Prostate Cancer Prostatic Dis. 2003;6(4):301-4.
- Chauhan D, Li G, Podar K, et al. A novel carbohydrate-based therapeutic GCS-100 overcomes bortezomib resistance and enhances dexamethasone-induced apoptosis in multiple myeloma cells. Cancer Res. 2005 Sep 15;65(18):8350-8.
- Johnson KD, Glinskii OV, Mossine VV, et al. Galectin-3 as a potential therapeutic target in tumors arising from malignant endothelia. Neoplasia. 2007 Aug;9(8):662-70.
- Streetly MJ, Maharaj L, Joel S, Schey SA, Gribben JG, Cotter FE. GCS-100, a novel galectin-3 antagonist, modulates MCL-1, NOXA, and cell cycle to induce myeloma cell death. Blood. 2010 May 13;115(19):3939-48.
- Wang Y, Nangia-Makker P, Balan V, Hogan V, Raz A. Calpain activation through galectin-3 inhibition sensitizes prostate cancer cells to cisplatin treatment. Cell Death Dis. 2010;1:e101.
- Tehranian N, Sepehri H, Mehdipour P, et al. Combination effect of PectaSol and Doxorubicin on viability, cell cycle arrest and apoptosis in DU-145 and LNCaP prostate cancer cell lines. Cell Biol Int. 2012 Jul;36(7):601-10.
- Jiang J, Eliaz I, Sliva D. Synergistic and additive effects of modified citrus pectin with two polybotanical compounds, in the suppression of invasive behavior of human breast and prostate cancer cells. Integr Cancer Ther. 2013 Mar;12(2):145-52.
- Yu L, Ruifrok WP, Meissner M, et al. Genetic and pharmacological inhibition of galectin-3 prevents cardiac remodeling by interfering with myocardial fibrogenesis. Circ Heart Fail. 2013 Jan;6(1):107-17.
- Liu YH, D’Ambrosio M, Liao TD, Peng H, Rhaleb NE, Sharma U, et al. N-acetyl-seryl-aspartyl-lysyl-proline prevents cardiac remodeling and dysfunction induced by galectin-3, a mammalian adhesion/growth-regulatory lectin. Am J Physiol Heart Circ Physiol. 2009 Feb;296(2):H404-12.
- Ahmad T, Felker GM. Galectin-3 in heart failure: more answers or more questions? J Am Heart Assoc. 2012 Oct;1(5):e004374.
- MacKinnon AC, Liu X, Hadoke PW, Miller MR, Newby DE, Sethi T. Inhibition of galectin-3 reduces atherosclerosis in apolipoprotein E-deficient mice. Glycobiology. 2013 Jun;23(6):654-63.
- Ahmad N, Gabius HJ, Sabesan S, Oscarson S, Brewer CF. Thermodynamic binding studies of bivalent oligosaccharides to galectin-1, galectin-3, and the carbohydrate recognition domain of galectin-3. Glycobiology. 2004 Sep;14(9):817-25.
- Liu FT, Hsu DK. The role of galectin-3 in promotion of the inflammatory response. Drug News Perspect. 2007 Sep;20(7):455-60.
- Ahmad N, Gabius HJ, André S, Kaltner H, Sabesan S, Roy R, Liu B, Macaluso F, Brewer CF. Galectin-3 precipitates as a pentamer with synthetic multivalent carbohydrates and forms heterogeneous cross-linked complexes. J Biol Chem. 2004 Mar 19;279(12):10841-7.
Wellness
Specialists
1-800-226-2370 - This service is FREE
7:30 AM - 12 AM (ET) Mon-Fri | 9 AM - 12 AM (ET) Sat-Sun