Life Extension Magazine®

Major Advance In Slowing Aging
A huge number of published studies demonstrate that higher AMPK activity can reverse degenerative aging processes by stimulating cellular energy metabolism, inhibiting fat storage, reducing triglyceride production, and boosting glucose uptake into muscles.
Scientifically reviewed by Dr. Gary Gonzalez, MD, in October 2024. Written by: William Faloon.

When we first described antioxidants in 1980, the concept was virtually unknown outside the scientific community.
Since then, the term “antioxidant” has become ubiquitous, as commercial companies widely use it to advertise products.
In this issue, we describe a method to counteract a deadly aging factor that is today as obscure as antioxidants were 35 years ago.
This novel approach is not new to most Life Extension® members. We have long recommended the drug metformin to protect against aging.1 The challenge in obtaining a doctor’s prescription has precluded many of our members from using metformin.
In a major advance, a pair of botanical extracts has been shown to work perhaps better than metformin to increase a critical enzyme (called AMPK) that enables cells to function with youthful vitality.
This article discusses the anti-aging effects that occur in response to higher AMPK activity. What should fascinate the public is a recent finding showing that ingestion of just one of these botanical extracts resulted in significant reductions of abdominal fat.2
Longevity enthusiasts will be thrilled to learn about the many degenerative processes that can be reversed when AMPK activity is enhanced.
The biological effects of increasing AMPK activity include inhibition of fat storage, reduced cholesterol-triglyceride synthesis, and increased glucose uptake into muscle.3-11 The diverse properties of AMPK may soon make this the most sought-after method to slow and reverse degenerative aging processes.
To understand the significance of AMPK, it is important to review some causes of aging that are not adequately addressed by the nutrients and hormones we currently take.
As we mature, our cells lose the ability to purge themselves of accumulated debris, often referred to as “cellular junk.”12-14 As junk-laden cells accumulate, they emit signals that generate low levels of inflammation.15 These senile cells lose their ability to degrade cellular junk through a process called autophagy. The accumulation of these defective “zombie” cells creates a catalyst for virtually all degenerative diseases.
The problems described in the previous paragraph can be traced to an AMPK deficit.15 Low AMPK activity cripples aging cells to the point where they no longer maintain their internal stability. This disturbance caused by insufficient AMPK signaling provokes disease and jeopardizes healthy aging.15
The term “energy” is a highly misused commercial term. Lay people associate it with caffeinated drinks that induce a temporary surge of adrenalin.
When we talk about enhancing energy metabolism through activation of AMPK, we are referring to turning back “on” youthful control of cellular functions that are critical to healthy longevity.
AMPK
stimulates energy metabolism by signaling cells to burn glucose and fatty acids.8,10,16-18
This is just one way that AMPK lowers blood glucose and reduces storage of body fat.
AMPK Declines With Aging

AMPK stands for adenosine monophosphate-activated protein kinase. It is an enzyme that plays a critical role in cellular homeostasis.19,20 AMPK acts as a master switch to regulate cell functions such as uptake of glucose, burning of fats, and formation of new mitochondria.15,21-25
When observing what happens to aging humans, such as steadily rising blood glucose levels and excess body fat accumulation, the impact of AMPK deficit can clearly be seen.
Nutritional overload impairs AMPK activity.26,27 It thus should not be surprising that one method of maintaining higher AMPK activity is calorie restriction.28,29 To better understand how this works, in a low-calorie environment, cells turn “on” survival signals (such as AMPK) to optimize their energy balance.30,31 Chronic overeating deactivates AMPK and shortens life spans.
Primate studies validate age-delaying effects when calorie restriction is properly executed.32-37 Humans find it challenging to consistently undereat. Fortunately, there are methods to mimic the AMPK-enhancing effects of low-calorie diets.
Cellular Housekeeping

Just imagine that you had a sawmill factory where debris was routinely removed as part of the manufacturing protocol. At some point, however, your workers decided to stop taking out the debris and wood chips start accumulating. This might not be an immediate problem, but as wood chip rubble piled up, your factory’s efficiency would decline, and at some point become dysfunctional.
Cells continuously produce metabolic waste products that are efficiently removed by AMPK signaling.15 As AMPK activity declines, waste products (cellular junk) accumulate and eventually render our cells dysfunctional.15
The dysfunction is so severe that senile cells cannot even commit suicide (apoptosis) because they lack sufficient energy instructions to perform even simple housekeeping tasks. Yet these senile cells emit chronic external signals that create a systemic state of low-level inflammation throughout the aging body.38-40
AMPK augments cellular housekeeping. Reduced AMPK signaling, on the other hand, can exacerbate common problems related to cellular dysfunction like heart failure.41 When you hear that an elderly person’s heart “wore out,” what often is being stated is that their cardiac muscle became dysfunctional due to reduced AMPK signaling.
Likewise, when type II diabetes manifests, it is often caused by a loss of cellular insulin sensitivity.42 AMPK improves insulin sensitivity,9,15 which is one mechanism by which the drug metformin lowers blood sugar levels.
AMPK Promotes Removal Of Senile Cells
Aging results in the accumulation of cells that can barely function yet create a host of problems such as chronic inflammation.
As AMPK activity declines with aging, defective cells linger and create metabolic havoc throughout the body.15,39,43,45 To purge the body of these senile cells, strong AMPK signaling44 is needed to facilitate the beneficial self-removal process.
Combatting Cellular Stress
Cellular stress occurs in response to extremes in temperature,46,47 exposure to toxins,48 mechanical damage,49 inflammation,50 and free radicals.51 A healthy stress response enables cells to survive these damaging/destructive events.
AMPK triggers signaling pathways that improve cell stress response.52
The ability of AMPK to react to cellular stress declines with age and this impairs the maintenance of cellular energy balance. In particular, a deficiency in AMPK signaling can make aging humans more vulnerable to the lethal impact of immune senescence and chronic inflammation.15
Boosting Sirtuin 1
The sirtuins are a family of genes involved in the regulation of cellular energy metabolism.
SIRT1 is one of the most studied of these genes because of its multifaceted role in cell survival, inflammation, and beneficial apoptosis (programmed cell death).15,53,54 SIR stands for “silent information regulator.”55
AMPK promotes the functional activity of SIRT1, which favorably influences beneficial longevity factors.56,57 Several studies indicate that SIRT1 signaling is associated with the extension of life span.58-60 SIRT1 increases during calorie restriction and can enhance cellular stress resistance, which is a well-known defense against the aging process.61-65
Resveratrol supplements have become popular because of their ability to enhance SIRT1.66-68 Resveratrol may do this by boosting AMPK.69 It is unlikely that resveratrol alone will optimally restore cellular AMPK activity to youthful ranges.
Controlling The Cell-Signaling Network
Researchers have identified several “signaling” pathways involved in the regulation of aging processes. These “signaling pathways” promote longevity in lower organisms.
One of these “signals,” named p53, controls cell proliferation.70 P53 is known as a tumor-suppressor gene and loss of p53 predisposes cells to malignancy.71-73 P53 is also involved in regulating cell metabolism and self-destruction (apoptosis) of senescent cells.74-76
Nuclear factor-kappa beta (NF-kB) is an internal cell signal that induces chronic inflammation.77,78 We take nutrients like curcumin to suppress NF-kB activation.79,80
By maintaining higher AMPK activity, our functional p53 is protected, while pro-inflammatory NF-kB is suppressed.
Summary Of AMPK
AMPK is a critical regulator of energy metabolism. The initial benefits of AMPK activation are lower glucose and triglyceride blood levels, along with reductions in abdominal fat mass.3-9,15,80-86
AMPK enables the desirable elimination of dysfunctional cell components (autophagy), which helps reduce chronic low-grade inflammation.87
Cellular stress resistance improves in response to greater AMPK activation.15
Increased AMPK activity can extend life span in lower organisms.15,88,89 The efficient clearance of “zombie” cells and improved cellular stress response are characteristics by which AMPK can enhance one’s ability to enjoy healthy longevity.
To state this simply, AMPK controls an integrated signaling network that has a major role in the regulation of the aging process.
How To Boost AMPK

Exercise activates AMPK, though this effect diminishes as one matures into their elder years.90,91 The ability of metformin to boost AMPK activity has caused some people to call metformin “exercise in a bottle.”92-95
In response to reduced calorie intake, cells activate AMPK96 as a survival mechanism.28,29 This beneficial AMPK activation vanishes when normal food consumption resumes.26,27
Nutrients like resveratrol and quercetin have some effect on activating AMPK, but are probably not as impactful as metformin.18,97 Those taking metformin are probably achieving optimal AMPK activation.
For those not aggressively exercising or taking at least 1,000 mg a day of metformin, a combination of two botanical extracts has emerged as perhaps the most effective way to activate cellular AMPK.98
The Animal Data
A 2011 published study on diabetic mice compared the effect of one of these botanical extracts to metformin. After 15 days, there was up to a 23% reduction in glucose in the
metformin-administered mice. The group receiving the botanical extract saw an up to 30% reduction in glucose.99
A 2012 published mouse study showed the following results when one of these botanical extracts were administered orally:100
- 10.3% reduction in sub-cutaneous belly fat (compared to control group), and
- 15.5% reduction in deep visceral fat (compared to control group).
The fat that builds up around the internal abdominal organs (called visceral fat) is the most dangerous form of body fat. Visceral fat generates chronic pro-inflammatory signals101 and distorts hormone balance.102-104 Most humans need to reduce their abdominal fat mass to avoid age-related disease.
The Human Data
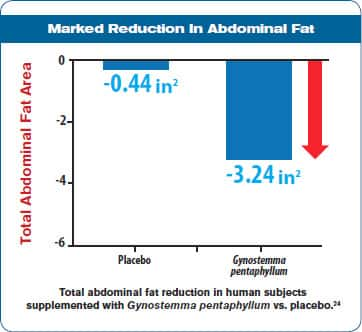
In 2014, a randomized, double-blind, placebo-controlled study was published that evaluated the effects of one of these AMPK-enhancing agents on body composition. Eighty human subjects were evaluated. One group received one of the new AMPK-augmenting botanical extracts and the second group received placebo.
The average body mass index (BMI) of the study subjects was 27.53, meaning they were clinically overweight but not obese. After 12 weeks, the group receiving the botanical extract showed reduction of 3.24 square inches in abdominal fat area (not waist circumference), whereas the placebo arm lost only 0.44 square inches.2 The botanical extract group lost more than an inch in abdominal circumference and nearly one-half inch in hip circumference, both of which are risk factors for the fat-driven inflammation that produces cardiovascular and metabolic diseases.
Reductions in belly fat have been reported in some people who take metformin, so it is not surprising that this novel botanical extract that activates cellular AMPK would reduce abdominal fat.
Fat accumulation in the abdomen105,106 and liver107-109 is often accompanied by the reduction in AMPK activity similar to what occurs during aging.15,110 For those who have been unable to shed meaningful weight in their bellies, these botanical extracts could provide the energy needed for abdominal cells to burn their surplus stored lipids (fat).
Decades Of Research
The first mention of AMPK on the National Library of Medicine’s database occurred in 1971.111
It was not until 2001 that published research linked decline in AMPK activity with pathological aging and shortened life spans.112
Life Extension® and others have spent enormous amounts of time and money investigating compounds that restore youthful AMPK activity. We long ago urged widespread use of metformin, but our members were challenged to find doctors to write prescriptions for this drug that the FDA approves only to treat type II diabetes.
With the discovery that two botanical extracts that have been safely used for centuries also promote AMPK activity, aging humans have a potent new weapon in their arsenal of age-delaying and potentially age-reversing compounds.
Annual Super Sale
In 1989, Life Extension® introduced a program that enables members to obtain advanced nutritional formulas at the year’s lowest prices.
By purchasing in quantities of four to 10 bottles, members stock up at steep discount prices on nutrients they need to stave off degenerative processes—such as decline in cellular AMPK activity.
In 2014, Life Extension® funded over $8.4 million in biomedical research aimed at finding cures for today’s common ailments and eradicating premature death. This research would not be possible if it were not for the loyalty members show every time they choose a Life Extension® supplement.
For longer life,
William Faloon
References
- Available at: http://www.medicalnewstoday.com/articles/280725.php?tw#citations. Accessed October 15, 2014.
- Park SH, Huh TL, Kim SY, et al. Antiobesity effect of Gynostemma pentaphyllum extract (actiponin): a randomized, double-blind, placebo-controlled trial. Obesity (Silver Spring). 2014 Jan;22(1):63-71.
- Villena JA, Viollet B, Andreelli F, Kahn A, Vaulont S, Sul HS. Induced adiposity and adipocyte hypertrophy in mice lacking the AMP-activated protein kinase-alpha2 subunit. Diabetes. 2004 Sep;53(9):2242-9.
- Anthony NM, Gaidhu MP, Ceddia RB. Regulation of visceral and subcutaneous adipocyte lipolysis by acute AICAR-induced AMPK activation. Obesity (Silver Spring). 2009 Jul;17(7):1312-7.
- Daval M, Diot-Dupuy F, Bazin R, et al. Anti-lipolytic action of AMP-activated protein kinase in rodent adipocytes. J Biol Chem. 2005 Jul 1;280(26):25250-7.
- Henin N, Vincent MF, Gruber HE, Van den Berghe G. Inhibition of fatty acid and cholesterol synthesis by stimulation of AMP-activated protein kinase. FASEB J. 1995 Apr;9(7):541-6.
- Henriksen BS, Curtis ME, Fillmore N, Cardon BR, Thomson DM, Hancock CR. The effects of chronic AMPK activation on hepatic triglyceride accumulation and glycerol 3-phosphate acyltransferase activity with high fat feeding. Diabetol Metab Syndr. 2013 May 31;5:29.
- Friedrichsen M, Mortensen B, Pehmoller C, Birk JB, Wojtaszewski JF. Exercise-induced AMPK activity in skeletal muscle: role in glucose uptake and insulin sensitivity. Mol Cell Endocrinol. 2013 Feb 25;366(2):204-14.
- Towler MC, Hardie DG. AMP-activated protein kinase in metabolic control and insulin signaling. Circ Res. 2007 Feb 16;100(3):328-41.
- Rutter GA, Da Silva Xavier G, Leclerc I. Roles of 5’-AMP-activated protein kinase (AMPK) in mammalian glucose homoeostasis. Biochem J. 2003 Oct 1;375(Pt 1):1-16.
- Pirkmajer S, Kulkarni SS, Tom RZ, et al.Methotrexate promotes glucose uptake and lipid oxidation in skeletal muscle via AMPK activation. Diabetes. 2014 Oct 22.
- Hoh Kam J, Lenassi E, Jeffery G. Viewing ageing eyes: diverse sites of amyloid Beta accumulation in the ageing mouse retina and the up-regulation of macrophages. PLoS One . 2010 Oct 1;5(10).
- Aprahamian T, Takemura Y, Goukassian D, Walsh K. Ageing is associated with diminished apoptotic cell clearance in vivo. Clin Exp Immunol. 2008 Jun;152(3):448-55.
- Neumann H, Kotter MR, Franklin RJ.Debris clearance by microglia: an essential link between degeneration and regeneration. Brain. 2009 Feb;132(Pt 2):288-95.
- Salminen A, Kaarniranta K. AMP-activated protein kinase (AMPK) controls the aging process via an integrated signaling network. Ageing Res Rev. 2012 Apr;11(2):230-41.
- Bijland S, Mancini SJ, Salt IP. Role of AMP-activated protein kinase in adipose tissue metabolism and inflammation. Clin Sci (Lond). 2013 Apr;124(8):491-507.
- Carling D. The AMP-activated protein kinase cascade—a unifying system for energy control. Trends Biochem Sci. 2004 Jan;29(1):18-24.
- Hardie DG. Sensing of energy and nutrients by AMP-activated protein kinase. Am J Clin Nutr. 2011 Apr;93(4):891S-6.
- Lage R, Diéguez C, Vidal-Puig A, López M. AMPK: a metabolic gauge regulating whole-body energy homeostasis. Trends Mol Med. 2008 Dec;14(12):539-49.
- Hardie DG, Ross FA, Hawley SA. AMPK: a nutrient and energy sensor that maintains energy homeostasis. Nat Rev Mol Cell Biol. 2012 Mar 22;13(4):251-62.
- Winder WW, Hardie DG. AMP-activated protein kinase, a metabolic master switch: possible roles in type 2 diabetes. Am J Physiol. 1999 Jul;277(1 Pt 1):E1-10.
- Andrade BM, Cazarin J, Zancan P, Carvalho DP. AMP-activated protein kinase upregulates glucose uptake in thyroid PCCL3 cells independent of thyrotropin. Thyroid. 2012 Oct;22(10):1063-8.
- Brusq JM, Ancellin N, Grondin P, et al. Inhibition of lipid synthesis through activation of AMP kinase: an additional mechanism for the hypolipidemic effects of berberine. J Lipid Res. 2006 Jun;47(6):1281-8.
- Reznick RM, Zong H, Li J, et al. Aging-associated reductions in AMP-activated protein kinase activity and mitochondrial biogenesis. Cell Metab. 2007 Feb;5(2):151-6.
- O’Neill HM, Maarbjerg SJ, Crane JD, et al. AMP-activated protein kinase (AMPK) beta1beta2 muscle null mice reveal an essential role for AMPK in maintaining mitochondrial content and glucose uptake during exercise. Proc Natl Acad Sci USA. 2011 Sep 20;108(38):16092-7.
- Coughlan KA, Valentine RJ, Ruderman NB, Saha AK. Nutrient excess in AMPK downregulation and insulin resistance. J Endocrinol Diabetes Obes. 2013; Sep 14.
- Saha AK, Xu XJ, Balon TW, Brandon A, Kraegen EW, Ruderman NB. Insulin resistance due to nutrient excess: is it a consequence of AMPK downregulation? Cell Cycle. 2011 Oct 15;10(20):3447-51.
- Lee SH, Min KJ. Caloric restriction and its mimetics. BMB Rep. 2013 Apr;46(4):181-7.
- Ribaric S. Diet and aging. Oxid Med Cell Longev. 2012;741468.
- Canto C, Auwerx J. Calorie restriction: is AMPK as a key sensor and effector? Physiology (Bethesda). 2011 Aug;26(4):214-24.
- Boily G , Seifert EL, Bevilacqua L, et al. SirT1 regulates energy metabolism and response to caloric restriction in mice. PLoS One. 2008 Mar 12;3(3):e1759.
- McKiernan SH, Colman RJ, Aiken E, et al. Anderson RM. Cellular adaptation contributes to calorie restriction-induced preservation of skeletal muscle in aged rhesus monkeys. Exp Gerontol . 2012 Mar;47(3):229-36.
- Kemnitz JW. Calorie restriction and aging in nonhuman primates. ILAR J. 2011 52(1):66-77.
- Colman RJ, Anderson RM, Johnson SC, et al. Caloric restriction delays disease onset and mortality in rhesus monkeys. Science. 2009 Jul 10;325(5937):201-4.
- Kastman EK, Willette AA, Coe CL, et al. A calorie-restricted diet decreases brain iron accumulation and preserves motor performance in old rhesus monkeys. J Neurosci. 2010 Jun 9;30(23):7940-7.
- Bodkin NL, Alexander TM, Ortmeyer HK, Johnson E, Hansen BC. Mortality and morbidity in laboratory-maintained Rhesus monkeys and effects of long-term dietary restriction. J Gerontol A Biol Sci Med Sci. 2003 Mar;58(3):212-9.
- Lane MA, Black A, Handy A, Tilmont EM, Ingram DK, Roth GS. Caloric restriction in primates. Ann N Y Acad Sci. 2001 Apr;928:287-95.
- Freund A, Orjalo AV, Desprez PY, Campisi J. Inflammatory networks during cellular senescence: causes and consequences. Trends Mol Med. 2010 16(5):238-46.
- Cevenini E, Monti D, Franceschi C. Inflamm-ageing. Curr Opin Clin Nutr Metab Care. 2013 Jan;16(1):14-20.
- Ren JL, Pan JS, Lu YP, Sun P, Han J. Inflammatory signaling and cellular senescence. Cell Signal. 2009 Mar;21(3):378-83.
- Turdi S, Fan X, Li J, et al. AMP-activated protein kinase deficiency exacerbates aging-induced myocardial contractile dysfunction. Aging Cell. 2010 Aug;9(4):592-606.
- Guilherme A, Virbasius JV, Puri V, Czech MP.Adipocyte dysfunctions linking obesity to insulin resistance and type 2 diabetes. Nat Rev Mol Cell Biol. 2008 May;9(5):367-77.
- Salminen A, Kaarniranta K. AMP-activated protein kinase (AMPK) controls the aging process via an integrated signaling network. Ageing Res Rev. 2012 Apr;11(2):230-41.
- Guo L, Xie B, Mao Z. Autophagy in Premature Senescent Cells Is Activated via AMPK Pathway. Int J Mol Sci. 2012;13(3):3563-82.
- Rubinsztein DC, Mariño G, Kroemer G. Autophagy and aging. Cell. 2011 Sep 2;146(5):682-95.
- Mosser DD, Caron AW, Bourget L, Denis-Larose C, Massie B.Role of the human heat shock protein hsp70 in protection against stress-induced apoptosis. Mol Cell Biol. 1997 Sep;17(9):5317-27.
- Ragoonanan V, Hubel A, Aksan A.Response of the cell membrane-cytoskeleton complex to osmotic and freeze/thaw stresses. Cryobiology. 2010 Dec;61(3):335-44.
- Tesh VL. Activation of cell stress response pathways by Shiga toxins. Cell Microbiol. 2012 Jan;14(1):1-9.
- Baskurt OK, Meiselman HJ. Red blood cell mechanical stability test. Clin Hemorheol Microcirc. 2013;55(1):55-62.
- Khansari N, Shakiba Y, Mahmoudi M.Chronic inflammation and oxidative stress as a major cause of age-related diseases and cancer. Recent Pat Inflamm Allergy Drug Discov. 2009 Jan;3(1):73-80.
- Rahman K. Studies on free radicals, antioxidants, and co-factors. Clin Interv Aging. 2007;2(2):219-36.
- Luo Z, Zang M, Guo W.AMPK as a metabolic tumor suppressor: control of metabolism and cell growth. Future Oncol. 2010 Mar;6(3):457-70.
- Hwang JW, Yao H, Caito S, Sundar IK, Rahman I. Redox regulation of SIRT1 in inflammation and cellular senescence. Free Radic Biol Med. 2013 Mar 27;61C:95-110.
- Yang H, Zhang W, Pan H, Feldser HG, Lainez E, Miller C, et al. SIRT1 activators suppress inflammatory responses through promotion of p65 deacetylation and inhibition of NF-κB activity. PLoS One. 2012 7(9):e46364.
- Hernández-Jiménez M, Hurtado O, Cuartero MI, Ballesteros I, Moraga A, Pradillo JM, McBurney MW, Lizasoain I, Moro MA.Silent information regulator 1 protects the brain against cerebral ischemic damage. Stroke. 2013 Aug;44(8):2333-7.
- Wang Y, Liang Y, Vanhoutte PM. SIRT1 and AMPK in regulating mammalian senescence: a critical review and a working model. FEBS Lett. 2011 Apr 6;585(7):986-94.
- Salminen A, Hyttinen JM, Kaarniranta K. AMP-activated protein kinase inhibits NF-κB signaling and inflammation: impact on health span and life span. J Mol Med (Berl). 2011 Jul;89(7):667-76.
- Longo VD, Kennedy BK. Sirtuins in aging and age-related disease. Cell. 2006 Jul 28;126(2):257-68.
- Cantó C, Auwerx J. Caloric restriction, SIRT1 and longevity. Trends Endocrinol Metab. 2009 Sep;20(7):325-31.
- Haigis MC, Sinclair DA. Mammalian sirtuins: biological insights and disease relevance. Annu Rev Pathol. 2010 5:253-95.
- Yu W, Zhou HF, Lin RB, Fu YC, Wang W. Short-term calorie restriction activates SIRT1 4 and 7 in cardiomyocytes in vivo and in vitro. Mol Med Rep. 2014 Apr;9(4):1218-24.
- Haigis MC, Guarente LP. Mammalian sirtuins--emerging roles in physiology, aging, and calorie restriction. Genes Dev. 2006 Nov 1;20(21):2913-21.
- Rogina B, Helfand SL. Sir2 mediates longevity in the fly through a pathway related to calorie restriction. Proc Natl Acad Sci U S A. 2004 Nov 9;101(45):15998-6003.
- Chen D, Bruno J, Easlon E, et al. Tissue-specific regulation of SIRT1 by calorie restriction. Genes Dev. 2008 Jul 1;22(13):1753-7.
- Cohen DE, Supinski AM, Bonkowski MS, Donmez G, Guarente LP. Neuronal SIRT1 regulates endocrine and behavioral responses to calorie restriction. Genes Dev. 2009 Dec 15;23(24):2812-7.
- Ghosh S, Liu B, Zhou Z. Resveratrol activates SIRT1 in a lamin A-dependent manner. Cell Cycle. 2013 Mar 15;12(6):872-6.
- Liu B, Zhou Z. Activation of SIRT1 by resveratrol requires lamin A. Aging (Albany NY). 2013 Feb;5(2):94-5.
- Lin CH, Lin CC, Ting WJ, et al. Resveratrol enhanced FOXO3 phosphorylation via synergetic activation of SIRT1 and PI3K/Akt signaling to improve the effects of exercise in elderly rat hearts. Age (Dordr). 2014 Oct;36(5):9705.
- Chen S, Xiao X, Feng X, et al. Resveratrol induces Sirt1-dependent apoptosis in 3T3-L1 preadipocytes by activating AMPK and suppressing AKT activity and survivin expression. J Nutr Biochem . 2012 Sep;23(9):1100-12.
- Duursma A, Agami R. p53-Dependent regulation of Cdc6 protein stability controls cellular proliferation. Mol Cell Biol. 2005 Aug;25(16):6937-47.
- Eischen CM, Weber JD, Roussel MF, Sherr CJ, Cleveland JL. Disruption of the ARF-Mdm2-p53 tumor suppressor pathway in Myc-induced lymphomagenesis. Genes Dev. 1999 Oct 15;13(20):2658-69.
- Chen Z, Trotman LC, Shaffer D, et al. Crucial role of p53-dependent cellular senescence in suppression of Pten-deficient tumorigenesis. Nature. 2005 Aug 4;436(7051):725-30.
- Ventura A, Kirsch DG, McLaughlin ME, et al. Restoration of p53 function leads to tumour regression in vivo. Nature. 2007 Feb 8;445(7128):661-5.
- Liang Y, Liu J, Feng Z. The regulation of cellular metabolism by tumor suppressor p53. Cell Biosci. 2013 Feb 6;3(1):9.
- Purvis JE, Karhohs KW, Mock C, Batchelor E, Loewer A, Lahav G. p53 dynamics control cell fate. Science. 2012 Jun 15;336(6087):1440-4.
- Lowe J, Shatz M, Resnick MA, Menendez D. Modulation of immune responses by the tumor suppressor p53. BioDiscovery. 2013;8:2.
- Hoesel B, Schmid JA. The complexity of NF-κB signaling in inflammation and cancer. Mol Cancer. 2013 Aug 2;12:86.
- Pal S, Bhattacharjee A, Ali A, Mandal NC, Mandal SC, Pal M. Chronic inflammation and cancer: potential chemoprevention through nuclear factor kappa B and p53 mutual antagonism. J Inflamm (Lond). 2014 Aug 9;11:23.
- Singh S , Aggarwal BB. Activation of transcription factor NF-kappa B is suppressed by curcumin (diferuloylmethane) [corrected]. J Biol Chem. 1995 Oct 20;270(42):24995-5000.
- Shishodia S, Amin HM, Lai R, Aggarwal BB. Curcumin (diferuloylmethane) inhibits constitutive NF-kappaB activation, induces G1/S arrest, suppresses proliferation, and induces apoptosis in mantle cell lymphoma. Biochem Pharmacol . 2005 Sep 1;70(5):700-13.
- Hardie DG. AMP-activated protein kinase: a master switch in glucose and lipid metabolism. Rev Endocr Metab Disord. 2004;5:119-25.
- Muoio DM, Seefeld K, Witters LA, Coleman RA. AMP-activated kinase reciprocally regulates triacylglycerol synthesis and fatty acid oxidation in liver and muscle: evidence that sn-glycerol-3-phosphate acyltransferase is a novel target. Biochem J. 1999 Mar 15;338(Pt 3):783-91.
- Murase T, Misawa K, Haramizu S, Minegishi Y, Hase T. Nootkatone, a characteristic constituent of grapefruit, stimulates energy metabolism and prevents diet-induced obesity by activating AMPK. Am J Physiol Endocrinol Metab. 2010 Aug;299(2):E266-75.
- Lee MS, Kim IH, Kim CT, Kim Y. Reduction of body weight by dietary garlic is associated with an increase in uncoupling protein mRNA expression and activation of AMP-activated protein kinase in diet-induced obese mice. J Nutr. 2011 Nov;141(11):1947-53.
- Park JE, Kim JY, Kim J, et al. Pomegranate vinegar beverage reduces visceral fat accumulation in association with AMPK activation in overweight women: A double, randomized, and placebo-controlled trial. J Funct Foods. 2014 May;(8):274-81.
- Cool B, Zinker B, Chiou W, et al. Identification and characterization of a small molecule AMPK activator that treats key components of type 2 diabetes and the metabolic syndrome. Cell Metab . 2006 Jun;3(6):403-16.
- Salminen A, Kaarniranta K, Kauppinen A. Inflammaging: disturbed interplay between autophagy and inflammasomes. Aging (Albany NY). 2012 Mar;4(3):166-75.
- Stenesen D, Suh JM, Seo J, et al. Adenosine nucleotide biosynthesis and AMPK regulate adult life span and mediate the longevity benefit of caloric restriction in flies. Cell Metab. 2013 Jan 8;17(1):101-12.
- Ulgherait M, Rana A, Rera M, Graniel J, Walker DW. AMPK modulates tissue and organismal aging in a non-cell-autonomous manner. Cell Rep. 2014 Sep 25;8(6):1767-80.
- Hardie DG. AMP-activated protein kinase: a key system mediating metabolic responses to exercise. Med Sci Sports Exerc. 2004 Jan;36(1):28-34.
- Mortensen B, Poulsen P, Wegner L, et al. Genetic and metabolic effects on skeletal muscle AMPK in young and older twins. Am J Physiol Endocrinol Metab. 2009 Oct;297(4):E956-64.
- Hawley SA, Gadalla AE, Olsen GS, Hardie DG. The antidiabetic drug metformin activates the AMP-activated protein kinase cascade via an adenine nucleotide-independent mechanism. Diabetes. 2002 Aug;51(8):2420-5.
- Matsui Y, Hirasawa Y, Sugiura T, Toyoshi T, Kyuki K, Ito M. Metformin reduces body weight gain and improves glucose intolerance in high-fat diet-fed C57BL/6J mice. Biol Pharm Bull. 2010 33(6):963-70.
- Kristensen JM, Treebak JT, Schjerling P, Goodyear L, Wojtaszewski JF. Two weeks of metformin treatment induces AMPK-dependent enhancement of insulin-stimulated glucose uptake in mouse soleus muscle. Am J Physiol Endocrinol Metab. 2014 May 15;306(10):E1099-109.
- Available at: http://www.bloomberg.com/news/2014-11-18/exercise-in-a-bottle-is-next-food-frontier-for-nestle.html. Accessed November 21, 2014.
- Palacios OM, Carmona JJ, Michan S, et al.Diet and exercise signals regulate SIRT3 and activate AMPK and PGC-1alpha in skeletal muscle. Aging (Albany NY). 2009 Aug 15;1(9):771-83.
- Hwang JT, Kwon DY, Yoon SH. AMP-activated protein kinase: a potential target for the diseases prevention by natural occurring polyphenols. N Biotechnol. 2009 Oct 1;26(1-2):17-22.
- Muller C, Gardemann A, Keilhoff G, Peter D, Wiswedel I, Schild L. Prevention of free fatty acid-induced lipid accumulation, oxidative stress, and cell death in primary hepatocyte cultures by a Gynostemma pentaphyllum extract. Phytomedicine. 2012 Mar 15;19(5):395-401.
- Qiao W, Zhao C, Qin N, Zhai HY, Duan HQ. Identification of trans-tiliroside as active principle with anti-hyperglycemic, antihyperlipidemic and antioxidant effects from Potentilla chinesis. J Ethnopharmacol. 2011 May 17;135(2):515-21.
- Gauhar R, Hwang SL, Jeong SS, et al. Heat-processed Gynostemma pentaphyllum extract improves obesity in ob/ob mice by activating AMP-activated protein kinase. Biotechnol Lett. 2012 Sep;34(9):1607-16.
- Item F, Konrad D. Visceral fat and metabolic inflammation: the portal theory revisited. Obes Rev. 2012 Dec;13 Suppl 2:30-9.
- Wisse BE. The inflammatory syndrome: the role of adipose tissue cytokines in metabolic disorders linked to obesity. J Am Soc Nephrol. 2004 Nov;15(11):2792-800.
- Nieves DJ, Cnop M, Retzlaff B, et al. The atherogenic lipoprotein profile associated with obesity and insulin resistance is largely attributable to intra-abdominal fat. Diabetes. 2003 Jan;52(1):172-9.
- Lim CT, Kola B, Korbonits M. AMPK as a mediator of hormonal signalling. J Mol Endocrinol. 2010 Feb;44(2):87-97.
- Gauthier MS, O’Brien EL, Bigornia S, et al. Decreased AMP-activated protein kinase activity is associated with increased inflammation in visceral adipose tissue and with whole-body insulin resistance in morbidly obese humans. Biochem Biophys Res Commun . 2011 Jan 7;404(1):382-7.
- Xu XJ, Gauthier MS, Hess DT, et al. Insulin sensitive and resistant obesity in humans: AMPK activity, oxidative stress, and depot-specific changes in gene expression in adipose tissue. J Lipid Res . 2012 Apr;53(4):792-801.
- Miyamoto L, Ebihara K, Kusakabe T, et al. Leptin activates hepatic 5’-AMP-activated protein kinase through sympathetic nervous system and α1-adrenergic receptor: a potential mechanism for improvement of fatty liver in lipodystrophy by leptin. J Biol Chem . 2012 Nov 23;287(48):40441-7.
- Lindholm CR, Ertel RL, Bauwens JD, Schmuck EG, Mulligan JD, Saupe KW. A high-fat diet decreases AMPK activity in multiple tissues in the absence of hyperglycemia or systemic inflammation in rats. J Physiol Biochem . 2013 Jun;69(2):165-75.
- García-Villafranca J, Guillén A, Castro J. Ethanol consumption impairs regulation of fatty acid metabolism by decreasing the activity of AMP-activated protein kinase in rat liver. Biochimie . 2008 Mar;90(3):460-6.
- Ix JH, Sharma K. Mechanisms linking obesity, chronic kidney disease, and fatty liver disease: the roles of fetuin-A, adiponectin, and AMPK. J Am Soc Nephrol. 2010 Mar;21(3):406-12.
- Eil C, Wool IG. Phosphorylation of rat liver ribosomal subunits: partial purification of two cyclic AMP activated protein kinases. Biochem Biophys Res Commun. 1971 Jun 4;43(5):1001-9.
- Tissenbaum HA, Guarente L. Increased dosage of a sir-2 gene extends life span in Caenorhabditis elegans. Nature. 2001 Mar 8;410(6825):227-30.
Wellness
Specialists
1-800-226-2370 - This service is FREE
7:30 AM - 12 AM (ET) Mon-Fri | 9 AM - 12 AM (ET) Sat-Sun