Life Extension Magazine®
One of the major building blocks of the brain, the omega-3 fatty acid docosahexaenoic acid (DHA) is critical for optimal brain health and function at all ages of life. Researchers are now finding that DHA provides brain-boosting benefits in infants and aging adults. Recognizing DHA’s crucial role in neurological and visual development in infants, manufacturers are rushing to incorporate DHA into infant formulas and baby foods. Similarly, adults seeking to stave off psychiatric and neurological ailments such as depression, post-traumatic stress disorder, and Alzheimer’s disease are now paying careful attention to their DHA intake.Here, we’ll discuss DHA’s integral role in ensuring optimal cognitive and neurological health. What you need to know
Multiple Benefits of Omega-3 OilsMost scientists and medical authorities agree that a higher intake of omega-3 oils is important for good health. This is based on a large number of well-conducted studies in humans and animals, which have shown that omega-3 oils improve brain function, decrease inflammation, reduce the incidence of fatal heart attacks and ischemic strokes, improve the outcome of autoimmune diseases, and improve vision. Of particular interest are the beneficial effects of omega-3 oils on neurological function and brain protection. For instance, a number of studies have shown that higher intakes of omega-3 oils significantly reduce the incidence of Alzheimer’s disease as well as vascular dementia, and improve quality of life and memory in those affected by dementia.1 Although there is general agreement about the health benefits of omega-3 oils, few appreciate the differences in their various components called eico-sapentaenoic acid (EPA) and docosahexaeonic acid (DHA). These two components make up the bulk of the omega-3 oils. The ratio of EPA and DHA can vary considerably in commercial products, however, most of which have much higher concentrations of EPA, mainly because it is less expensive to manufacture and has a less fishy odor than DHA. Yet, studies have shown that most of the neurological benefits of omega-3 oils are derived from the DHA component rather than the EPA component.
In both the Chicago2 and Rotterdam studies,3 researchers found a 60% reduction in the incidence of Alzheimer’s disease in those eating at least one fish-containing meal a week. In the Chicago study, DHA exerted a strong protective effect, which was not seen with EPA. Furthermore, this study found that intake of the plant-derived omega-3 fatty acid, alpha-linolenic acid, was associated with a reduction in the risk of Alzheimer’s disease in subjects carrying the APOE4 gene, which is a powerful indicator of the disease. People with Alzheimer’s disease have dramatically lower levels of DHA in the neurons of their hippocampus, an area of the brain severely affected in the disease. This area is vital to creating recent memories (so-called working memory or declarative memory).4 Prasad and co-workers from the University of Kentucky found that phospholipids such as phosphatidyl-ethanolamine (PE), which normally contain the highest levels of DHA, are severely depleted in those regions of the brain most affected by Alzheimer’s disease.5
DHA supplementation not only improves memory in cases of Alzheimer’s disease but has also been shown to improve age-related memory loss as well.6 Animal studies have shown that DHA-supplemented diets can counteract learning difficulties when soluble amyloid-beta (the most injurious form of Alzheimer’s-associated amyloid) is infused into the animals’ brains.7 Animal studies, both in mice and non-human primates, also show that DHA-depleted diets impair learning and memory, and that re-feeding DHA-containing diets reverses these impairments.8-9 Feeding rats diets high in DHA improves both working memory (very short-term memory) and reference memory (longer-term) in both old and young animals.8,10,11 Reisbick and co-workers at the Oregon Health Sciences University found that rhesus monkeys fed a long-term omega-3-deficient diet began to show stereotyped behaviors during early life.12 These are the type of repetitive behaviors seen with social deprivation and autism. Clearly, omega-3 oils such as DHA play essential roles in learning, memory, and behavior. Next, we’ll look at their role in supporting nervous system development. DHA and Brain DevelopmentA growing number of studies in both human infants and animals are showing a strong correlation between the intake of omega-3 oils, especially DHA, and cognitive function, visual acuity, and overall brain development.13-15 In the human infant, brain development undergoes its most rapid and complex growth during the last trimester of pregnancy and the first two years after birth. This means that during this period the child’s eventual neurological development is highly dependent on its dietary intake of essential nutrients, especially omega-3 oils. Because the fetal and infant brain is unable to convert enough alpha-linolenic oil (an omega-3 oil found in some plant oils) into DHA, the baby almost totally depends on its mother for its supply.16 There is good evidence that the placenta selectively takes up DHA so as to ensure an adequate supply for the growing baby.17,18 Some 70% of energy supply during fetal development is devoted to brain development, and lipids make up 50 to 60% of the structure of the brain, with DHA making up 30% of the brain and 50% of the retina’s structure. However, since the baby depends on the mother for its supply of DHA, omega-3 deficiencies in the mother can lead to increases in DHA deficiency in the infant brain with each successive birth. Studies have shown that brain DHA deficiency is limited to about 30% with the first generation, but by the third generation, brain DHA levels can fall by as much as 85%.19 The average DHA and EPA intake combined in the USA is about 100-200 mg/day, far below the 650 mg of DHA plus EPA that experts recommend for healthy individuals.20 Furthermore, it has been shown that after birth, DHA levels fall between the ages of 6 and 12 months mostly due to low DHA content in most baby foods and formulas,21 which can have a profound effect on postnatal brain development. A study that measured the impact of DHA on visual development in 6-month-old infants found that retinal and visual cortex maturation was greatly improved in infants receiving DHA-enriched egg yolk versus control infants who were deficient in DHA. In fact, the researchers found that even in breast-fed infants, DHA levels fell, whereas levels rose by 34% in the supplemented infants.21 In developing babies, higher levels of DHA levels are needed for the growth of neuronal cells than other brain structures.22 The composition of the membranes of neural cells is under constant reconstruction, and can change drastically with changes in diet. It has been shown that while other fatty acids plateau at birth, DHA levels actually double, mainly during periods of intense nerve cell development, that is, during intense brain growth. Overwhelming evidence for the benefits of DHA comes from a recent review of some 50 studies, which concluded that higher DHA in babies’ diets translates into better brain function, especially for cognitive and visual function.23 Further evidence that DHA may promote healthy nervous system development comes from a recent study in which researchers divided 53 normal, healthy infants into those exclusively breast-fed and those exclusively bottle-fed with a formula containing no DHA. Using assessments of visual, auditory, and sensory perception, they found definite abnormalities in all three tests in the bottle-fed babies at one year of age.24 A devastating consequence of low DHA status is its impact on the development of neurological abnormalities. In another study, researchers measured DHA, essential fatty acids, trans fatty acids, and arachidonic acid levels in the umbilical artery and vein of infants. They found that infants with neurological abnormalities at birth had significantly lower levels of arachidonic acid and DHA and higher trans fatty acid levels.25 In contrast, infants with higher arachidonic acid, DHA, and essential fatty acid levels had more normal neurological function. Editor’s note: While excess levels of arachidonic acid contribute to chronic inflammation in aging humans, arachidonic acid is a critical building block to developing brains. In a further test of cognitive development in infancy, researchers found that maternal and fetal DHA levels measured at birth correlated with attention and distractibility at 12 and 18 months, with those having higher DHA levels fairing better than those with low levels.26 It is also interesting to note that DHA plays a major role in muscle function and coordination in developing infants. A recent study using newborn baboons revealed that the highest levels of DHA are found in the motor control areas of the brain,27 which also play a major role in memory, social development, and behavior. So we see that adequate DHA in the mother’s diet is critical for normal brain development in her babies, and that with the first baby, her DHA level will fall even more because the placenta will preferentially extract the mother’s DHA for the baby. This means that subsequent babies will have even lower DHA levels as the mother’s DHA is further depleted. The big question is, can developmental problems triggered by low DHA levels be reversed? The evidence seems to indicate, yes. For example, in one study, rats fed low-DHA diets for three generations from birth (producing severe deficiencies in brain DHA levels) and then fed a DHA-enhanced diet for six weeks achieved the same level on testing of spatial learning as animals maintained on an omega-3-adequate diet for three generations.28 A DHA-enriched diet for less than six weeks had no effect. However, the rate of recovery of brain DHA varies among studies, depending on the experimental animal model and other variables. In another study, rat pups that were fed DHA-depleted diets for two generations followed by an omega-3-containing diet beginning at 15 days after birth began to regain DHA in essential brain structures within one day, but full recovery took one month.29 This slow recovery in rats was confirmed by other scientists.30 In the rhesus monkey, recovery of brain DHA can take as long as 12 weeks.31 In addition, the brain recovers its lost DHA slower than peripheral tissues. In one study, the rate of recovery of brain DHA went from 19% at one week, to 35% at two weeks, and to 80% at eight weeks in rats. In the retina, which has an even higher DHA content than the brain, recovery was faster than the brain, reaching 72% at four weeks, and was fully recovered at eight weeks.32 Recovery of DHA by the adult brain, however, is slower and less complete than the infant brain.33 DHA and the Adult BrainWhile DHA is essential for the proper development of the infant brain, it also plays a vital role in the ongoing structure and function of the adult brain. It is a component of several important phospholipids in the brain, with the highest levels of DHA being found in phosphatidylethanolamine (PE) and phosphatidyl-serine (PS).34,35 Lower levels exist in phosphatidyl-cholines (PC). Structurally, DHA comprises 22 carbons and six double bonds, making it the most unsaturated fatty acid in cell membranes and an important ingredient in increasing the fluidity of cell membranes. Changing the fluidity of cell membranes alters their physical properties, such as permeability and protein activity. This change can drastically alter cell signaling and plays a major role in increasing membrane permeability of cancer cells, making them more susceptible to immunologic and chemothera-peutic killing.36 EPA, however, has much less effect on membrane fluidity than DHA.37 It has also been shown that a greater incorporation of DHA into cell membranes results in dramatically less susceptibility to lipid peroxidation and oxidative stress in cells, especially neurons.38 Although many view the brain as a stable structure that changes little after adolescence, it is in fact always in a state of flux, not only by forming millions of new nerve connections, but also by replacing and altering its biochemical makeup, especially membrane lipids. Connected with this constant turnover of brain lipids are the phospholipase enzymes, which can release arachidonic acid and DHA from the cell membrane.39 Dietary changes can therefore drastically alter brain lipids, which can significantly alter brain function, even worsening neuropsychiatric disorders.40 Furthermore, there is evidence that as we age, the distribution of DHA in the brain changes.41 For example, during infancy the highest levels are found in the striatum (associated with motor control) and are lower in the hypothalamus (linking the nervous and endocrine systems) and hippocampus (associated with memory). In adults, the highest levels are in the cortex (essential to cognition) and lowest in the medulla (crucial for autonomic function). With aging, the highest levels are in the cortex and cerebellum (involved in motor control). An example of this regional specificity was seen in a recent study when scientists restored dietary DHA to rats that had been deprived of the nutrient. They found that all areas of brain DHA were restored after 12 weeks, except for the medulla, which recovered only 62% of its DHA.41 These findings therefore provide plentiful evidence that consuming enough DHA may be essential for upkeep of the adult brain. DHA and Cell SignalingIn the past, a lot of attention has been paid to the effects of omega-3 oils on membrane properties, which are very important to the brain’s healthy functioning. After all, membranes regulate entry into the cell as well as control receptor function, which facilitates cellular communication. More recently, however, researchers have discovered that omega-3 oils, especially the DHA component, also affect cell signaling.
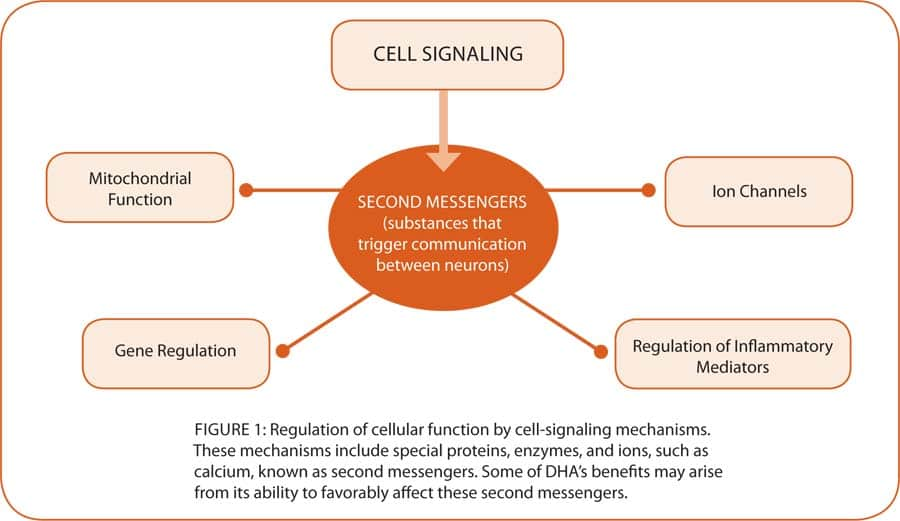
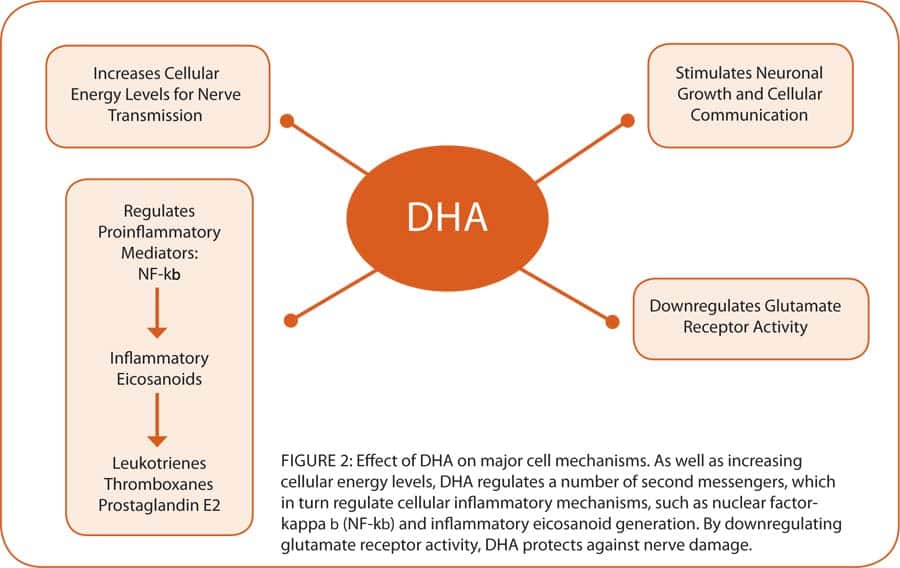
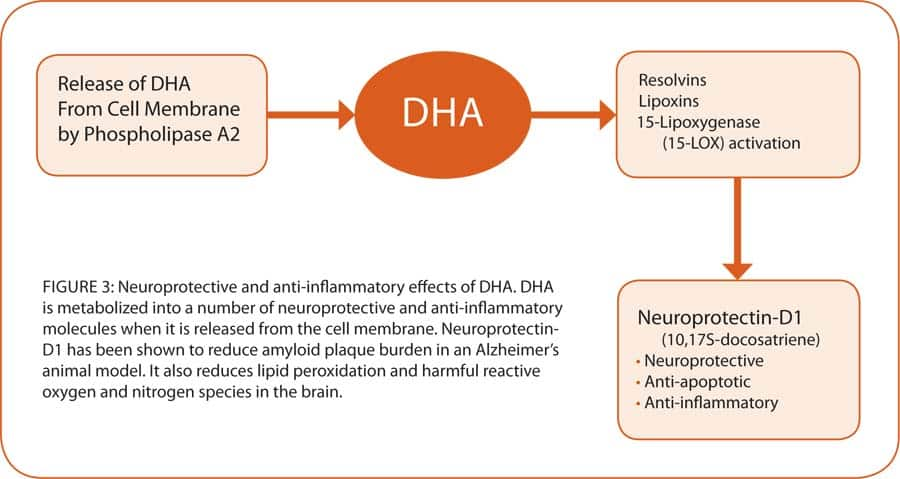
Hundreds, if not thousands, of special molecules are involved in cell signaling, which control not only the cell’s internal communication with the outside, but also communication within the cell. A group of special transcription molecules allows cells to communicate with the genes as well. The vast array of cell-signaling molecules allows the signals to be fine-tuned and alternately switched to an assortment of functions—such as regulating cellular energy via the mitochondria, activating or silencing genes, generating specialized proteins, ion regulation, and regulating inflammatory mediators (see figure 1 on page 50). The complexity of actions within a cell is therefore carefully regulated by cell-signaling molecules. Numerous enzymes, such as adenyl cyclase and protein kinase A involved in energy regulation, are upregulated by DHA. The DHA omega-3 also reduces proinflammatory mediators such as prostaglandin E2, thromboxanes, and leukotrienes, and increases the production of anti-inflammatory compounds such as lipoxins and resolvin along with substances that protect brain cells called neuroprotectins (see figure 3 on page 51).42
DHA also maintains the activity of a key cell membrane enzyme called Na+/K+ ATPase that extracts energy from ATP to drive the cellular sodium pump, which controls electrical impulses betweens cells. For example, it has been shown that feeding rats a DHA-deficient diet lowers activity of Na+/K+ ATPase by 40%.43 This enzyme is also linked to 60% of the energy consumed by the brain.44 Maintaining cellular energy levels is particularly important in preventing excitotoxicity (a pathological process that damages or kills nerve cells), since low energy can dramatically increase sensitivity to this destructive process. Another essential cognitive process controlled by DHA is regulating the brain’s concentration of phosphatidylserine (PS), which is also vital for cell survival.45 Diets low in DHA reduce brain PS, which affects cell signaling for survival through enzymes including Na+/K+ ATPase and calcium uptake. Calcium is one of the most common and versatile cell-signaling systems. DHA, but not EPA, has been shown to be a major regulator of calcium oscillations (waves within cells),46 which regulate a vast array of cellular functions, including neurotransmitter release, mitochondrial function, gene activation, oxidative stress, and, in the developing brain, neuron migration and maturation.47 Low DHA levels are also known to lower brain and cellular growth factors, such as brain-derived growth factor. Another detrimental effect of low DHA is to increase neuronal death. An observation of this was seen in a lab study, which revealed a dramatic increase in DNA fragmentation of DHA-depleted neurons in cell culture. Enriching this cellular medium with DHA protected the neurons,48 an effect that appeared to operate through phosphatidylserine (PS) via its actions on cell-signaling survival pathways.
As well as killing neurons, diets low in DHA decrease PS levels in the hippocampal region of the brain, associated with memory consolidation, which may explain, in part, the improvement in cognition and memory seen in studies of PS supplementation in patients with dementia.49 DHA is the major omega-3 oil component of the central nervous system and accumulates particularly in PS. Omega-3 deficiency has been correlated with approximately 30-35% reduction in PS levels in key neuronal cells during both prenatal and postnatal periods.96 One of the principal cell-signaling pathways affected by both EPA and DHA is the peroxisome proliferator-activated receptors, or PPARs. When activated, this pathway inhibits proinflammatory cytokine production, thus greatly reducing inflammation, both systemically and in the brain. This could play a major role in reducing the damage generated by a host of disorders such as diabetes, atherosclerosis, autoimmune diseases, brain aging, autism, and the neuro-degenerative disorders. Studies have shown that EPA and DHA activate PPARs, which in turn suppress nuclear factor-kappab (NF-kb), the main transcription system that activates inflammation.50 Thus, DHA, which is more potent than EPA in activating PPARs, may have important implications for reducing inflammation in the brain. A high intake of DHA and EPA also reduces inflammation by displacing arachidonic acid (a proinflammatory precursor) and cholesterol from the cell membrane, reducing the starting material used to make inflammatory mediators. Furthermore, DHA inhibits inflammatory eicosanoid production induced by the release of arachidonic acid from cell membranes. As we age, brain inflammation progressively increases, but a diet higher in DHA relative to other fatty acids may reduce this age-related brain inflammation. |
|||||||||



DHA and NeurotransmittersThe beneficial effects of DHA extend beyond promoting optimal neuron function and reducing inflammation. A number of studies have shown that omega-3 oils—especially DHA—can also support healthy levels of neurotransmitters, the chemical messengers between cells. For example, it has been shown that a DHA-supplemented diet can restore neurotransmitter release in the hippocampus and reverse age-related impairment of memory acquisition.51 This may be due, in part, to DHA’s effect on increasing the fluidity of cell membranes, which improves their ability to release neurotransmitters, but it may also involve cell signaling.52,53 Another important effect of DHA is in increasing the density of nerve outgrowths called dendrites, a property that is strongly connected to improved learning in rats as well as humans.54
In addition, DHA has rather complex effects on a number of neurotransmitters, including serotonin, norepinephrine, acetylcholine, glutamate, and dopamine.46,55,56 Deficiencies in DHA have been shown to lower the level of dopamine and the dopamine receptor D2 in the frontal lobe, which can result in problems with attention and learning.57 This may explain the ability of DHA to promote focused attention and to calm anxiety. One important effect of DHA-deficient diets was seen in a study using the reward chemical dopamine, which found that long-term omega-3 deficiency raised dopamine levels in a region of the brain closely linked to addiction behavior.58 This, in essence, significantly increases susceptibility to addictive behaviors, whether to foods, sex, or illicit drugs. Finally, DHA also plays a key role in neurotrans-mitter receptor function. Recent studies have found significant effects of DHA on cell membrane structures called G-protein-coupled receptors, which communicate with the inside of the cell when stimulated by neurotransmitters such as glutamate, serotonin, and acetylcholine.42,59 Since these receptors are inducible, that is, their numbers can be increased on demand or by altering nutrients, dietary changes can significantly affect how the brain functions. Effect of Omega-3 Oils on Depression and Suicide RiskBecause of the frequent profound effect of EPA, and especially DHA, on cell signaling, membranes, neurotransmitters and receptors, one would expect that omega-3 oils produce equally profound changes in human behavior as well. In fact, a number of studies have found a close correlation between omega-3 oils in the diet and behavior. Of special interest is the effect of omega-3 oils on depression, suicide risk, aggression, anger, and anxiety.
Depression has become a major problem, with rates increasing at younger ages than ever before, and the disease occurring even in preteens. Also increasing are rates of anxiety disorders, anger dyscontrol, insomnia, violent suicides, and even homicides. Scientists have proposed a common link between these disorders and chronic brain inflammation. Likewise, a number of medical conditions have been associated with chronic depression, such as cardiovascular disease, cancer, and overall poor health. Until recently, it was assumed that this was because of neuro-endocrine effects or other nonspecific reasons. However, a growing amount of research now indicates these diseases are linked to depression by a common mechanism—a high level of inflam-matory mediators, such as cytokines and inflam-matory prostaglandins. Unlike acute brain inflammation, as seen with meningitis and encephalitis, subtle, chronic brain inflammation can be silent, except for its behavioral manifestations.60 Chronic brain inflammation can be the result of a number of conditions, such as abdominal obesity, latent viral infections, autoimmune diseases, metabolic syndrome, heavy metal intoxi-cation, and deficiencies in omega-3 oils. Studies have revealed a close correlation between chronic inflammation and depression.61-65 A recent study even found a strong correlation between inflammatory cytokine levels and the high rates of depression and suicide in anesthesiologists.66 Furthermore, a number of studies suggest that many of the antidepressant medications also act as anti-inflammatory agents and that patients who respond poorly have the highest levels of inflammatory cytokines.67-70 Chronic brain inflammation has also been found in post-traumatic stress disorder and postpartum depression.71,72 It may be that omega-3 oils improve major depression, as well as a number of other neuro-psychiatric disorders, by reducing brain inflammation. It is known that a deficiency in omega-3 oils affects serotonergic and dopaminergic neuro-transmission, and that if restored before weaning (discontinuation of nursing), these defects can be repaired in the brain.73,74 Both of these neurotransmitters, as well as inflammation, play a significant role in neuro-psychiatric disorders such as schizophrenia, borderline psychotic personality, suicide, hostility/aggression, bipolar disorder, and major depression. Shrinkage of the hippocampus of the brain has been described in major depression and in post-traumatic stress disorder.75 This shrinkage (atrophy) is also accompanied by deficiencies in recent memory. Memory deficits may occur before atrophy and neuron loss, and treatment of the depression may help restore hippocampal neurons and connections (neutrites and synapses).76
While both EPA and DHA have been associated with improvements in major depression, only DHA stimulates neurite outgrowth and synaptic development and repair, that is, brain plasticity.54 Therefore, it may be a better choice in methods to repair the brain damage of chronic depression. A growing problem in this country is postpartum depression, which is often linked with suicide, homicide, and infanticide. A number of studies have linked low blood levels and red blood cell levels of omega-3 fatty acids with postpartum depression.77-79 For example, a study of 10 pregnant women who developed postpartum depression versus 38 who did not found that the proportion of DHA in the serum phospholipids was consistently lower in the women with postpartum depression.79 Suicide risk has also been linked to low omega-3 levels in the blood and cell membranes.80,81 In one such study, researchers at the Department of Neuroscience at the New York State Psychiatric Institute followed 33 medication-free, depressed patients for two years. Seven of the subjects attempted suicide. Low plasma DHA percentage in serum phospholipids and a high ratio of omega-6 fats to omega-3 fats accurately predicted the suicide attempts.82 Unfortunately, Americans consume far more omega-6 fats than are needed for good health. With the aforementioned effects of DHA on brain levels of the neurotransmitters serotonin and dopamine, as well as receptor expression and cell signaling in synaptic membranes, one should not be surprised by the dramatic behavioral impact of omega-3 deficiency. The evidence for the effect of omega-3 oils on another neurotransmitter, glutamate—the most abundant messenger in the brain—is still being debated. There is compelling evidence that a number of neuropsychiatric disorders, including schizophrenia, biopolar disorder, suicide risk, and major depression, are related to abnormalities in glutamate neurotransmission.83-85 Data to indicate that excess glutamate may be responsible for major depression, as well as other psychiatric disorders of mood and impulse control comes from a number of quarters. Firstly, depression and schizophrenia have been linked with a decrease in glutamate transport proteins, which normally carry excess glutamate away from nerve endings. This can greatly increase brain glutamate levels and excitotoxicity,86 and would explain the loss of neurons in the hippocampus seen in schizophrenia and major depression. Secondly, the drug ketamine, which is a powerful glutamate receptor antagonist, also acts like an antidepressant.87 Another drug, riluzole, which prevents the release of glutamate from cells also prevents degeneration of brain cells in depression and raises brain growth factor levels, which are consistently lowered in depression. Finally, careful studies have shown that glutamate receptors in depression are overactivated and glutamate removal systems, which normally protect the brain from glutamate excess, function abnormally. These same studies have found that the gamma-aminobutyric acid (GABA) system, which is essential to protecting the brain against excitotoxicity, also works subnormally. If indeed, major depression, bipolar disorder, schizophrenia, and suicide risk are related to excitotoxic injury, omega-3 oils may play a significant role in combating these disabling disorders, since several studies have shown that omega-3 oils—especially DHA—significantly reduce excitotoxicity.88,89 Low DHA in the Diet Increases Anger and HostilityLow omega-3 levels, especially DHA levels, have also been associated with anger and hostility reactions. For example, in the first of a two-part multicenter randomized study, medical students were either given a placebo or fish oils containing 1.5 g per day of DHA for three months.90 In this part of the study, which was done during a high-stress period (during final exams), they found an 8.9% increase in aggression in the placebo group, whereas those taking the DHA experienced no increase in aggressive tendencies. In the second part of the study, using 46 non-stressed students, DHA had no effect on aggression.91 This is because increases in aggression were secondary to stress and DHA appeared to only calm stimulated aggressive urges. This is consistent with the above-cited studies indicating DHA deficiency causes disruption of monoamine neurotransmission (dopamine, norepinephrine, serotonin) as well as acetylcholine and glutamate neurotransmission, all of which are related to impulse and anger control. These effects are also related to the cell-signaling, anti-inflammatory, and membrane fluidity effects of DHA, which are greater than those of EPA (see figure 3 on page 51). In a later study in 41 university students, researchers compared a high-DHA diet (1.5 g/day) with placebo (soybean oil) over a three-month period to see the effects on hostile responses.92 They found that there was a 27% incidence of hostile answers at baseline, which rose to 92% during final exams. Again, there was no increase in hostility among those taking the high-DHA supplement. They also found that DHA supplementation significantly reduced hostility measures among university staff. There is even evidence that extra DHA in the diet can reduce aggressive behavior in younger children. For example, in a study examining girls and boys aged 9 to 12 years, adding omega-3-fortified foods (3,600 mg DHA and 840 mg EPA per week) reduced impulsive behavior in both males and females, with stronger results in females.93 One of the effects of higher DHA intake is to reduce plasma norepinephrine levels, which reduces the fight-or-flight response. In one study, DHA’s ability to modulate plasma norepinephrine was quite dramatic, with no effects on cortisol, epinephrine, glucose, or insulin levels.94 DHA could thus be a useful weapon to reduce anxiety and panic attacks. Supplementing with DHAHealthy adults should consume 240-1,000 mg of DHA daily to protect their cognitive and neurological health. While many adults already consume omega-3 oil supplements containing both DHA and EPA, DHA standalone products allow those seeking higher DHA/EPA ratios to customize their intake according to their unique health concerns. Pregnant and lactating women require a minimum of 300 mg DHA daily to promote optimal nervous system development in their children. Parents should ensure that their young children consume at least 150 mg of DHA per day to support health. Omega-3 oils are also available in good-tasting liquid formulations for young children and those who have difficulty swallowing capsules. SummaryFrom this review we have seen that the omega-3 oils play a major role in brain development, with the highest demand occurring during intrauterine development and the first two years after birth (during the brain growth spurt). Deficiencies during neuro-development in the mother alter not only immediate development, but can set the stage for later disease and brain malfunction. Because of the high demand for DHA by the developing baby and the preferential extraction from the mother’s supply by the fetus, deficiencies are compounded with succeeding births, should the diet not improve. Likewise, this has a profound effect on the psychological profile of the mother, with severe deficiencies of her DHA resulting in increased rates of postpartum depression, and concomitant increased risk of suicide and homicide. We have also seen that the effects of DHA and EPA on nervous system cells go beyond structural membrane effects (fluidity), but rather influence cell signaling, neurotransmitters, and receptor expression and function. This also includes gene regulation and expression. The effects of omega-3 oils on inflammation are complex, and go far beyond physically displacing arachidonic acid—the starting material for inflammatory prostaglandins—from the cell membrane. DHA also increases lipoxins, inhibits NF-kb, and produces neuroprotectin-D1 and resolvins. Because of the complex interaction of omega-3 oils with a multitude of systems, both cellular and via neuronal networks, variations in brain EPA and especially DHA can have profound effects on learning and behavior. Ensuring optimal amounts of these healthful oils is thus essential throughout the healthy life span. Safety CautionIndividuals who use the medication warfarin (Coumadin®) should consult a physician before supplementing with DHA.95 If you have any questions on the scientific content of this article, please call a Life Extension Health Advisor at 1-800-226-2370. | |||
| References | |||
| 1. Yehuda S, Rabinovtz S, Carasso RL, Mostofsky DI. Essential fatty acids preparation (SR-3) 2. Morris MC, Evans DA, Bienias JL, et al. Consumption of fish and omega-3 fatty acids and risk of incident 3. Kalmijn S, Launer LJ, Ott A, Witteman JC, Hofman A, Breteler MM. 4. Soderberg M, Edlund C, Kristensson K, Dallner G. Fatty acid composition of brain phospholipids in 5. Prasad MR, Lovell MA, Yatin M, Dhillon H, Markesbery WR. Regional membrane phospholipid 6. Morris MC, Evans DA, Tangney CC, Bienias JL, Wilson RS. Fish consumption and cognitive decline with age in a 7. Hashimoto M, Hossain S, Shimada T, et al. Docosahexaenoic acid provides protection from impairment of learning 8. Gamoh S, Hashimoto M, Hossain S, Masumura S. Chronic administration of docosahexaenoic acid improves the performance 9. Neuringer M, Connor WE. omega-3 fatty acids in the brain and retina: evidence for their essentiality. 10. Sugimoto Y, Taga C, Nishiga M, et al. Effect of docosahexaenoic acid-fortified Chlorella vulgaris strain CK22 on 11 Gamoh S, Hashimoto M, Sugioka K, et al. Chronic administration of docosahexaenoic acid improves reference 12. Reisbick S, Neuringer M, Hasnain R, Connor WE. Home cage behavior of rhesus monkeys 13. Enslen M, Milon H, Malnoe A. Effect of low intake of omega-3 fatty acids during development on brain phospholipid 14. Birch EE, Garfield S, Castaneda Y, et al. Visual acuity and cognitive outcomes at 4 years of age in a double-blind, 15. Willatts P, Forsyth JS, DiModugno MK, Varma S, Colvin M. Effect of long-chain polyunsaturated fatty acids 16. Cunnane SC, Williams SC, Bell JD, et al. Utilization of uniformly labeled 13C-polyunsaturated fatty acids in the synthesis of 17. Crabtree JT, Gordon MJ, Campbell FM, Dutta-Roy AK. Differential distribution and metabolism of arachidonic acid 18. Crawford MA, Hassam AG, Stevens PA. Essential fatty acid requirements in pregnancy and lactation with 19. Ozias MK, Carlson SE, Levant B. Maternal parity and diet (omega-3) polyunsaturated fatty acid concentration influence accretion of brain 20. Kris-Etherton PM, Taylor DS, Yu-Poth S, et al. Polyunsaturated fatty acids in the food chain in the United States. 21. Hoffman DR, Theuer RC, Castaneda YS, et al. Maturation of visual acuity is accelerated in breast-fed term infants fed baby food containing 22. Green P, Glozman S, Kamensky B, Yavin E. Developmental changes in rat brain membrane lipids and fatty acids. 23. McCann JC, Ames BN. Is docosahexaenoic acid, an omega-3 long-chain polyunsaturated fatty acid, required for 24. Khedr EM, Farghaly WM, Amry S, Osman AA. Neural maturation of breastfed and formula-fed infants. 25. Dijck-Brouwer DA, Hadders-Algra M, Bouwstra H, et al. Lower fetal status of docosahexaenoic acid, arachidonic acid 26. Colombo J, Kannass KN, Shaddy DJ, et al. Maternal DHA and the development of attention in infancy and toddlerhood. 27. Diau GY, Hsieh AT, Sarkadi-Nagy EA, et al. The influence of long chain polyunsaturate supplementation on docosahexaenoic 28. Moriguchi T, Salem N, Jr. Recovery of brain docosahexaenoate leads to recovery of spatial task performance. 29. Youyou A, Durand G, Pascal G, et al. Recovery of altered fatty acid composition induced by a diet devoid of omega-3 fatty acids in myelin, 30. Homayoun P, Durand G, Pascal G, Bourre JM. Alteration in fatty acid composition of adult rat brain capillaries and choroid plexus induced 31. Connor WE, Neuringer M, Lin DS. Dietary effects on brain fatty acid composition: the reversibility of omega-3 fatty acid deficiency and 32. Moriguchi T, Loewke J, Garrison M, Catalan JN, Salem N, Jr. Reversal of docosahexaenoic acid deficiency in the rat brain, retina, liver, and 33. Anderson GJ. Developmental sensitivity of the brain to dietary omega-3 fatty acids. J Lipid Res. 1994 Jan;35(1):105-11. 34. Stillwell W, Shaikh SR, Zerouga M, Siddiqui R, Wassall SR. Docosahexaenoic acid affects cell signaling by altering lipid rafts. Reprod 35. Stillwell W, Wassall SR. Docosahexaenoic acid: membrane properties of a unique fatty acid. Chem Phys Lipids. 2003 Nov;126(1):1-27. 36. Stillwell W, Ehringer W, Jenski LJ. Docosahexaenoic acid increases permeability of lipid vesicles and tumor cells. Lipids. 1993 Feb;28(2):103-8. 37. Brown ER, Subbaiah PV. Differential effects of eicosapentaenoic acid and docosahexaenoic acid on human skin fibroblasts. 38. Zerouga M, Jenski LJ, Stillwell W. Comparison of phosphatidylcholines containing one or two docosahexaenoic acyl chains on properties of 39. Horrobin DF, Bennett CN. New gene targets related to schizophrenia and other psychiatric disorders: enzymes, binding proteins and transport 40. Horrobin DF. Interactions between lipid metabolism and schizophrenia: the biochemical changes which may have made us human. Lipids. 41. Xiao Y, Huang Y, Chen ZY. Distribution, depletion and recovery of docosahexaenoic acid are region-specific in rat brain. 42. Litman BJ, Niu SL, Polozova A, Mitchell DC. The role of docosahexaenoic acid containing phospholipids in modulating G protein-coupled signaling 43. Bourre JM, Francois M, Youyou A, et al. The effects of dietary alpha-linolenic acid on the composition of nerve membranes, enzymatic activity, 44. Turner N, Else PL, Hulbert AJ. Docosahexaenoic acid (DHA) content of membranes determines molecular activity of 45. Salem N, Jr., Litman B, Kim HY, Gawrisch K. Mechanisms of action of docosahexaenoic acid in the nervous system. Lipids. 46. Chalon S, ion-Vancassel S, Belzung C, et al. Dietary fish oil affects monoaminergic neurotransmission and behavior in rats. 47. Sergeeva M, Strokin M, Reiser G. Regulation of intracellular calcium levels by polyunsaturated fatty acids, arachidonic acid and docosahexaenoic acid, 48. Kim HY, Akbar M, Lau A, Edsall L. Inhibition of neuronal apoptosis by docosahexaenoic acid (22:6omega-3). Role of phosphatidylserine in antiapoptotic effect. 49. Delwaide PJ, Gyselynck-Mambourg AM, Hurlet A, Ylieff M. Double-blind randomized controlled study of phosphatidylserine in senile demented patients. 50. Zhao G, Etherton TD, Martin KR et al. Anti-inflammatory effects of polyunsaturated fatty acids in THP-1 cells. 51. McGahon BM, Martin DS, Horrobin DF, Lynch MA. Age-related changes in synaptic function: analysis of the effect of dietary supplementation 52. Fujita S, Ikegaya Y, Nishikawa M, Nishiyama N, Matsuki N. Docosahexaenoic acid improves long-term potentiation attenuated by phospholipase 53. Kitajka K, Puskas LG, Zvara A, et al. The role of omega-3 polyunsaturated fatty acids in brain: modulation of rat brain gene expression by dietary 54. Cao D, Xue R, Xu J, Liu Z. Effects of docosahexaenoic acid on the survival and neurite outgrowth of rat cortical neurons in primary cultures. 55. de la Presa OS, Innis SM. Docosahexaenoic and arachidonic acid prevent a decrease in dopaminergic and serotoninergic neurotransmitters in 56. Innis SM, de la Presa OS. Dietary fatty acid composition in pregnancy alters neurite membrane fatty acids and dopamine in newborn rat brain. 57. Delion S, Chalon S, Guilloteau D, Besnard JC, Durand G. alpha-Linolenic acid dietary deficiency alters age-related changes of dopaminergic and 58. Zimmer L, Vancassel S, Cantagrel S, et al. The dopamine mesocorticolimbic pathway is affected by deficiency in omega-3 polyunsaturated fatty acids. 59. Mitchell DC, Niu SL, Litman BJ. Enhancement of G protein-coupled signaling by DHA phospholipids. Lipids. 2003 Apr;38(4):437-43. 60. Blaylock RL. Chronic microglial activation and excitotoxicity secondary to excessive immune stimulation: 61. Yang K, Xie G, Zhang Z, et al. Levels of serum interleukin (IL)-6, IL-1beta, tumour necrosis factor-alpha and leptin and their 62. Kim YK, Na KS, Shin KH, et al. Cytokine imbalance in the pathophysiology of major depressive disorder. 63. Kiecolt-Glaser JK, Belury MA, Porter K, et al. Depressive symptoms, omega-6:omega-3 fatty acids, and inflammation in older adults. 64. Craddock D, Thomas A. Cytokines and late-life depression. Essent Psychopharmacol. 2006;7(1):42-52. 65. Spalletta G, Bossu P, Ciaramella A, et al. The etiology of poststroke depression: a review of the literature and a new 66. Beilin B, Greenfeld K, Abiri N, et al. Anesthesiologists at work: an increase in pro-inflammatory and Th2 cytokine production, 67. O’Brien SM, Scully P, Fitzgerald P, Scott LV, Dinan TG. Plasma cytokine profiles in depressed patients who fail to 68. Basterzi AD, Aydemir C, Kisa C, et al. IL-6 levels decrease with SSRI treatment in patients with major depression. 69. O’Brien SM, Scott LV, Dinan TG. Antidepressant therapy and C-reactive protein levels. Br J Psychiatry. 2006 May;188:449-52. 70. Narita K, Murata T, Takahashi T, et al. Plasma levels of adiponectin and tumor necrosis factor-alpha in patients 71. von KR, Hepp U, Kraemer B, et al. Evidence for low-grade systemic proinflammatory activity in patients with posttraumatic stress disorder. 72. Levant B, Ozias MK, Carlson SE. Specific brain regions of female rats are differentially depleted of docosahexaenoic acid by 73. Kodas E, Galineau L, Bodard S, et al. Serotoninergic neurotransmission is affected by omega-3 polyunsaturated fatty acids in the rat. 74. Kodas E, Vancassel S, Lejeune B, Guilloteau D, Chalon S. Reversibility of omega-3 fatty acid deficiency-induced changes in 75. Hickie I, Naismith S, Ward PB, et al. Reduced hippocampal volumes and memory loss in patients with early- and late-onset depression. 76. MacMaster FP, Kusumakar V. Hippocampal volume in early onset depression. BMC Med. 2004 Jan 29;22. 77. Freeman MP. Omega-3 fatty acids and perinatal depression: a review of the literature and recommendations for future research. 78. Sontrop J, Campbell MK. Omega-3 polyunsaturated fatty acids and depression: a review of the evidence and a methodological critique. 79. De Vriese SR, Christophe AB, Maes M. Lowered serum omega-3 polyunsaturated fatty acid (PUFA) levels predict the occurrence 80. Huan M, Hamazaki K, Sun Y, et al. Suicide attempt and omega-3 fatty acid levels in red blood cells: a case control study in 81. Terao T, Soya A. Cholesterol, essential fatty acids, and suicide. Pharmacopsychiatry. 2003 Mar;36(2):86-7. 82. Sublette ME, Hibbeln JR, Galfalvy H, Oquendo MA, Mann JJ. Omega-3 polyunsaturated essential fatty acid status as a 83. Auer DP, Putz B, Kraft E, et al. Reduced glutamate in the anterior cingulate cortex in depression: an in vivo proton magnetic resonance 84. McCullumsmith RE, Kristiansen LV, Beneyto M, et al. Decreased NR1, NR2A, and SAP102 transcript expression in the hippocampus in 85. Noga JT, Hyde TM, Herman MM, et al. Glutamate receptors in the postmortem striatum of schizophrenic, suicide, and control brains. 86. McCullumsmith RE, Meador-Woodruff JH. Striatal excitatory amino acid transporter transcript expression in schizophrenia, 87. Mathew SJ, Keegan K, Smith L. Glutamate modulators as novel interventions for mood disorders. Rev Bras Psiquiatr. 2005 Sep;27(3):243-8. 88. Relton JK, Strijbos PJ, Cooper AL, Rothwell NJ. Dietary N-3 fatty acids inhibit ischaemic and excitotoxic brain damage in the rat. 89. Hogyes E, Nyakas C, Kiliaan A, et al. Neuroprotective effect of developmental docosahexaenoic acid supplement against excitotoxic 90. Hamazaki T, Sawazaki S, Nagasawa T, et al. Administration of docosahexaenoic acid influences behavior and plasma catecholamine 91. Hamazaki T, Sawazaki S, Nagao Y, et al. Docosahexaenoic acid does not affect aggression of normal volunteers under nonstressful conditions. 92. Hamazaki T, Sawazaki S, Itomura M, et al. Effect of docosahexaenoic acid on hostility. World Rev Nutr Diet. 2001;88:47-52. 93. Itomura M, Hamazaki K, Sawazaki S, et al. The effect of fish oil on physical aggression in schoolchildren—a randomized, double-blind, 94. Sawazaki S, Hamazaki T, Yazawa K, Kobayashi M. The effect of docosahexaenoic acid on plasma catecholamine concentrations and 95. Available at: http://www.pdrhealth.com/drugs/altmed/printview/altmed-mono-print.aspx?contentFileName=ame0278. 96. Hamilton L, Greiner R, Salem N Jr, Kim HY. n-3 fatty acid deficiency decreases phosphatidylserine accumulation selectively 97. Kelley DS, Siegel D, Vemuri M, Mackey BE. Docosahexaenoic acid supplementation improves fasting and postprandial lipid profiles in |



Wellness
Specialists
1-800-226-2370 - This service is FREE
7:30 AM - 12 AM (ET) Mon-Fri | 9 AM - 12 AM (ET) Sat-Sun