Life Extension Magazine®
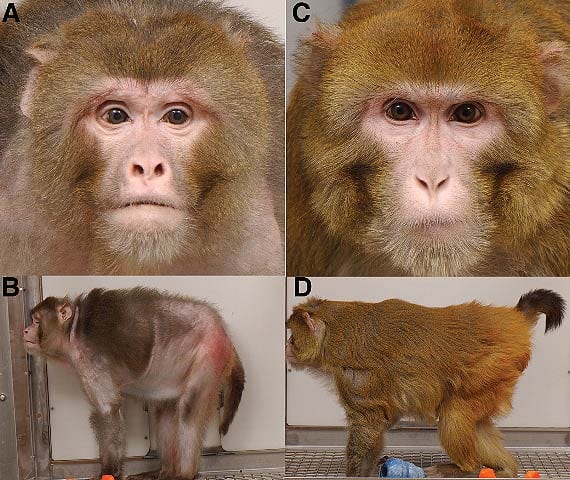
The most scientifically validated way to extend life span—from single-celled organisms to mammals—is caloric restriction.1 This technique has been shown to boost life span by nearly 100% in some species.2 Reducing the number of ingested calories—while maintaining healthy intake of essential nutrients—triggers a cascade of anti-aging mechanisms in the body. More than 70 years of research have established its life-extending power.3 Modern science has only recently begun to unravel the mystery behind how calorie restriction so radically extends life span. At its core lies favorable alterations in gene expression—one of the body’s adaptive responses to reduced calorie intake. By activating certain genes and disabling others, caloric restriction dramatically slows aging. This retardation of aging delays the onset of degenerative disease while improving biomarkers of youth, ranging from metabolic rate and insulin sensitivity to cardiac health and cognitive function.4 By the same token, the consequences of caloric overconsumption are equally profound. Every excess calorie brings you one step closer to age-related illness and death. This is true at any stage of life, whether you’re lean or overweight, regardless of your health or nutritional status.5-7 Eating more than your body needs (excessive energy intake) can load the blood with triglycerides, glucose, homocysteine, and pro-inflammatory chemicals. The results are accelerated aging processes, prompting deterioration across multiple biomarkers of health.8 Most humans find it difficult to submit to a sufficiently rigorous dietary regimen, leaving the benefits of calorie restriction tantalizingly out of reach. Until now! In this article you will discover the most compelling evidence to date on the effects of caloric restriction. In a milestone 20-year study of Rhesus monkeys—our close genetic relatives—a modestly restricted diet resulted in a three-fold reduction in the risk of age-related disease!9 You will also learn about five natural compounds that favorably modulate gene expression to support a more youthful health profile. These “caloric restriction mimics” simulate many of the beneficial effects of caloric restriction through multiple pathways. Aging individuals may now enjoy some of the youth-promoting, disease-fighting mechanisms of caloric restriction—without strict dietary measures. A Milestone in Longevity ResearchIn 1989, a group of anti-aging researchers launched what would prove to be the most comprehensive study of caloric restriction (CR) to date.10 Rhesus monkeys were chosen as subjects because they exhibit biological and aging characteristics strikingly similar to humans. The results, published in 2009 in the prestigious journal Science,9 provide compelling evidence of CR’s age-delaying, disease-fighting power. The monkeys were split into two groups. Half were allowed to eat naturally, without restraint; the other half subsisted on a nutrient-replete diet 30% lower in total calories than they would normally consume. After 20 years, 37% of controls had died of age-related causes, as opposed to just 13% in the calorie-restricted group. In other words, caloric restriction cut degenerative disease risk by a factor of three.9 This study’s findings are even more tantalizing when one sees that it did not require severe lowering of calorie intake to produce these striking results. Americans typically consume 100% more calories than they need. It only required a calorie reduction of 30% to achieve the remarkable benefits in this study. The Rhesus monkey study also conclusively demonstrated caloric restriction’s protective power. Over the course of 20 years, virtually all measured biomarkers of health were superior in the CR group.9 These findings have profound implications for humans. Rhesus monkeys are as vulnerable to chronic, age-related disease as we are. They lose their strength over time. Many become obese, fall prey to metabolic syndrome, and succumb to diabetes. They develop cardiovascular disease and cancer. And like aging humans, their brains shrink as they get older. Yet among the calorie-restricted group, incidence of cardiovascular disease was half the rate ofcontrols. Not one member exhibited any symptoms of impaired glucose control or diabetes, whereas 40% of monkeys who ate as much as they wanted had become diabetic or pre-diabetic.9 Calorie-restricted monkeys lost fat weight, but did not sustain loss of muscle mass observed in the control group. CR also inhibited reduction in brain volume, especially in areas governing cognitive and motor function.9 The robust health of the CR group compared to controls is clearly evident in photographs of the monkeys from the control and CR groups.9 (See figure 1 below.) Benefits of Calorie Restriction in Humans
All available data indicate that calorie restriction also slows aging and reduces the risk of killer diseases in humans as well. Individuals who impose a 20% reduction in their calorie consumption for 2-6 years lose fat weight and show significant improvement in their markers of aging, including blood pressure, cholesterol levels, and glucose control.11 Even brief periods of caloric restriction can temporarily improve core body temperature and insulin sensitivity, which happen to be markers of longevity.11,12 In clinical studies, short intervals of caloric restriction have reduced systemic inflammation—an underlying factor of so many chronic, degenerative diseases. More youthful heart muscle performance13 has also been observed—caloric restriction appears to increase the number of vital energy-producing mitochondria in heart and skeletal muscle, reducing the oxidative damage that accelerates aging.14-16 No other intervention documented in the scientific literature can compete with caloric restriction as a candidate for slowing aging and supporting youthful health in humans. It is regrettable that most aging individuals don’t lower calorie intake by the 20-30% reduction required to reap the benefits of caloric restriction, as it can dramatically lower disease risk and add years to their lives. Caloric Restriction “Mimics” and Gene ExpressionThe problem many people have in sufficiently reducing their calorie intake has led to a novel scientific solution. Researchers have identified a select group of nutrients that trigger many of the same underlying mechanisms of action as caloric restriction. Among the most promising of these caloric restriction mimics and enhancers are resveratrol, pterostilbene, quercetin, and grape seed extract, along with black tea extract. These nutraceuticals have been shown to generate many of the same effects in the body as caloric restriction, without significant dietary modification. In particular, they “mimic” caloric restriction’s favorable impact on genes that influence the aging process. Genes have the capacity to directly affect life span by regulating a broad spectrum of aging factors, from inflammation and metabolic function to immune response. Calorie restriction exerts a beneficial effect on the activity of gene expression, supporting healthy cellular function through numerous physiological pathways. This includes:
Compounds that mimic caloric restriction bring about favorable changes in gene expression and improve the primary biomarkers of aging.
Controlling Nuclear Factor-Kappa BCalorie restriction sharply limits expression of nuclear factor-kappa B (NF-kB). In the cells, NF-kB is a critical gene regulator that governs response to pro-inflammatory cytokines, free radicals, cholesterol levels, immune function, and cancer prevention.17-24 The nutrients that mimic caloric restriction also act to stabilize NF-kB regulation and help combat the age-related conditions associated with unfavorable NF-kB activity. Resveratrol activates sirtuins,25 a powerful family of “information regulator” proteins that inhibit NF-kB, reducing inflammation throughout the body, such as that caused by second-hand cigarette smoke.24 Resveratrol also prevents inflammatory mast cells from releasing the histamines that trigger asthma and allergic reactions.26 Resveratrol radically decreases production of the adhesion molecules that attract inflammatory cells to vascular walls, one of the principal mechanisms of atherosclerosis.27,28 Adhesion molecules also permit cancer cells to invade tissue and metastasize. Resveratrol’s influence over NF-kB has also enhanced beneficial UV-induced programmed cell death (apoptosis) in skin cancer cells.29 Found in blueberries, pterostilbene is a polyphenol closely related to resveratrol.30 It limits NF-kB activity through multiple complementary mechanisms.31 In vitro, pterostilbene suppresses invasive tumor activity and enhances therapeutic destruction of cancer cells.32,33 Quercetin’s ability to protect against chronic inflammatory conditions such as asthma, inflammatory bowel disease, and arthritis is due in part to its capacity for NF-kB inhibition.34,35 Grape seed extract also disrupts cellular inflammation signaling by blocking NF-kB.36 Its effect on pro-inflammatory cytokine production in fat cells may even help combat obesity and type 2 diabetes.37 Black tea extract’s influence on NF-kB activity has been shown to specifically protect cells from damage associated with inflammation,38 cancer,39,40 Parkinson’s disease,41 and stroke.42 Suppressing Inflammatory CytokinesCaloric restriction inhibits expression of genes that produce inflammatory cytokines—such as interleukins and tumor necrosis factor (TNF), as well as cyclooxygenase-2 (COX-2)—that are powerfully implicated in the onset of cancer, atherosclerosis, and chronic inflammation.43-46 Resveratrol and pterostilbene block the release of a host of inflammatory cytokines and enzymes found throughout the body—particularly tissues and organs stressed by environmental factors, infection, or trauma.22,47,48 Pterostilbene powerfully suppresses the expression of inflammatory COX-2,31,49 helping lower the risk of cancer as well as a host of inflammatory diseases.50,51 Quercetin inhibits the COX-2 enzyme involved in early-stage colon cancer, and suppresses numerous cytokines involved in allergies and autoimmune disorders.34,35,53 Grape seed extract specifically prevents fatty tissue from releasing inflammatory cytokines and adipokines that would otherwise provoke insulin resistance and atherosclerosis associated with metabolic syndrome.37 By downregulating reactive cytokines, grape seed extract has been shown in animals to protect organs from ischemia-reperfusion damage (injury caused by the return of blood flow after a period of restriction).54 Black tea extract reduces levels of STAT-1, a protein which “tells” the cell nucleus to activate genes that produce inflammatory cytokines.55 | ||||||

Cancer PreventionCalorie restriction (CR) upregulates genes that suppress cancer and downregulates genes that permit cancers to form or spread. CR prevents cancer cell reproduction and proliferation, while inhibiting the blood vessel growth cancer cells require to develop and metastasize.46,56-62
Resveratrol and pterostilbene mirror these effects. They combat cancer at every stage of development, inducing apoptosis (programmed cell death) in a variety of human cancer types, while preserving healthy cells.32,63-66 Resveratrol also suppresses cancer proliferation by modulating expression of proteins involved in the reproductive cycle of abnormal cells.67,68 Quercetin activates “executioner” proteins while inhibiting survival proteins in human cancer cells, blocking their reproduction.69-71 Quercetin and resveratrol have also been shown to block the expression of vascular endothelial growth factor (VEGF), an effect that may help starve tumors of their blood supply.72 Grape seed extract induces expression of a protein that arrests cancer cells early in their reproductive cycle, preventing further development and destroying them.73 Similar to quercetin, grape seed extract fights angiogenesis by suppressing the VEGF signaling pathway.74 Black tea extract reduces expression of genes that cancer cells use to proliferate, survive, infiltrate healthy tissue, supply themselves with blood, and metastasize to other organs.75 It has also been shown to upregulate expression of proteins that arrest the cell reproductive cycle and induce cellular death specifically in cancers.76 Enhanced Glucose ControlCaloric restriction enhances glucose control.77,78 Recall that not one of the Rhesus monkeys in the study discussed earlier developed diabetes or exhibited symptoms of impaired glucose control.9 Caloric restriction triggers gene regulators called peroxisome proliferator-activated receptors (PPARs), a class of proteins responsible for healthy fat and carbohydrate metabolism. They also play key roles in optimizing mitochondrial health57,59 and thwarting the onset of metabolic syndrome and diabetes.79 Resveratrol80 and pterostilbene81,82 upregulate the production and activity of PPAR, launching a set of cellular processes that support a youthful metabolic profile. The PPAR activator resveratrol has been shown to:
Grape seed extract modulates a different set of PPARs that regulate fat storage. Grape seed extract induces fat metabolism while inhibiting the development of new fat cells.86.87 It also protects endothelial cells by preventing the inflammatory response to proteins damaged by glucose (the age-accelerating process known as glycation).88 Resveratrol further exerts a favorable influence on blood sugar metabolism at the cellular level, reducing glucose production in liver cells in a way that mimics prolonged calorie restriction.89 In diabetic animals, resveratrol has been shown to help restore blood sugar to normal by modulating the activity of several enzymes involved in sugar metabolism.90 Pterostilbene and grape seed extract generate similar beneficial changes that help promote healthy blood sugar levels.91,92 Grape seed extract activates genes that trigger glucose uptake. This assists cells in the absorption and removal of glucose from circulation.92 Quercetin has been shown to stimulate the proliferation of pancreatic cells that help modulate blood glucose levels in both diabetic and non-diabetic animal models.93 It also markedly reduces expression of the enzyme that produces sorbitol, a sugar alcohol known to cause cataracts and blindess.94 Black tea extract polyphenols inhibit lipase,95 an enzyme that breaks down fat in the stomach and small intestines. This helps block absorption of fat into the bloodstream.96 In animal models, the theaflavins in black tea extract prevent after-meal elevations in blood glucose and may protect against the metabolic syndrome.97,98 This effect may help increase signaling for a powerful longevity factor called FOXO1a.99 SummaryCaloric restriction (CR) is the most scientifically validated method shown to reliably extend life span in multiple species, from microorganisms to mammals. A milestone 20-year study provides the most conclusive evidence to date of its efficacy in Rhesus monkeys, our close genetic relatives. The discovery of calorie restriction-mimicking nutrients makes it possible for aging humans to emulate some of CR’s beneficial mechanisms of action, especially as an adjunct to modestly reducing one’s overall calorie intake.100 The unique ability of these nutrients to modulate gene expression exerts system-wide effects that, in addition to influencing many of the same pathways activated by calorie restriction, can also significantly reduce degenerative disease risk. If you have any questions on the scientific content of this article, please call a Life Extension® Wellness Specialist at 1-866-864-3027. Editor's NoteScience continues to evolve, and new research is published daily. As such, we have a more recent article on this topic: Nutrients that Provide Benefits of Caloric Restriction | |
| References | |
| 1. Genaro Pde S, Sarkis KS, Martini LA. Effect of caloric restriction on longevity. Arq Bras Endocrinol Metabol. 2009 Jul;53(5):667-72. 2. McCay CM, Crowel MF. The Scientific Monthly. 1934 Nov;39(5):405–14. 3. McCay CM, Crowell MF, Maynard LA. The effect of retarded growth upon the length of life span and upon the ultimate body size. Nutrition. 1935;5:155-71. 4. Heilbronn L, Ravussin E. Calorie restriction and aging: review of the literature and implications for studies in humans. Am J Clin Nutr. 2003 Sept;78(3):361-9. 5. Fontana L, Villareal DT, Weiss EP, et al. Calorie restriction or exercise: effects on coronary heart disease risk factors. A randomized, controlled trial. Am J Physiol Endocrinol Metab. 2007 Jul;293(1):E197-202. 6. Tan CY, Vidal-Puig A. Adipose tissue expandability: the metabolic problems of obesity may arise from the inability to become more obese. Biochem Soc Trans. 2008 Oct;36(Pt 5):935-40. 7. Korner J, Woods SC, Woodworth KA. Regulation of energy homeostasis and health consequences in obesity. Am J Med. 2009 Apr;122(4 Suppl 1):S12-8. 8. Fontana L. Nutrition, adiposity and health. Epidemiol Prev. 2007 Sep-Oct;31(5):290-4. 9. Colman RJ, Anderson RM, Johnson SC, et al. Caloric restriction delays disease onset and mortality in rhesus monkeys. Science. 2009 Jul 10;325(5937):201-4. 10. Colman RJ, Roecker EB, Ramsey JJ, Kemnitz JW. The effect of dietary restriction on body composition in adult male and female rhesus macaques. Aging (Milano). 1998 Apr;10(2):83-92. 11. Everitt AV, Le Couteur DG. Life extension by calorie restriction in humans. Ann N Y Acad Sci. 2007 Oct;1114:428-33. 12. Heilbronn LK, de Jonge L, Frisard MI, et al. Effect of 6-month calorie restriction on biomarkers of longevity, metabolic adaptation, and oxidative stress in overweight individuals: a randomized controlled trial. JAMA. 2006 Apr 5;295(13):1539-48. 13. Holloszy JO, Fontana L. Caloric restriction in humans. Exp Gerontol. Aug 2007;42(8):709-12. 14. Menshikova EV, Ritov VB, Toledo FG, Ferrell RE, Goodpaster BH, Kelley DE. Effects of weight loss and physical activity on skeletal muscle mitochondrial function in obesity. Am J Physiol Endocrinol Metab. 2005 Apr;288(4):E818-25. 15. Civitarese AE, Carling S, Heilbronn LK, et al. Calorie restriction increases muscle mitochondrial biogenesis in healthy humans. PLoS Med. 2007 Mar;4(3):e76. 16. Heilbronn LK, Gan SK, Turner N, Campbell LV, Chisholm DJ. Markers of mitochondrial biogenesis and metabolism are lower in overweight and obese insulin-resistant subjects. J Clin Endocrinol Metab. 2007 Apr;92(4):1467-73. 17. Zou Y, Yoon S, Jung KJ, et al. Upregulation of aortic adhesion molecules during aging. J Gerontol A Biol Sci Med Sci. 2006 Mar;61(3):232-44. 18. Jung KJ, Maruyama N, Ishigami A, Yu BP, Chung HY. The redox-sensitive DNA binding sites responsible for age-related downregulation of SMP30 by ERK pathway and reversal by calorie restriction. Antioxid Redox Signal. 2006 Mar-Apr;8(3-4):671-80. 19. Kim HJ, Yu BP, Chung HY. Molecular exploration of age-related NF-kB/IKK downregulation by calorie restriction in rat kidney. Free Radic Biol Med. 2002 May 15;32(10):991-1005. 20. Chandrasekar B, Nelson JF, Colston JT, Freeman GL. Calorie restriction attenuates inflammatory responses to myocardial ischemia-reperfusion injury. Am J Physiol Heart Circ Physiol. 2001 May;280(5):H2094-102. 21. Chung HY, Kim HJ, Kim KW, Choi JS, Yu BP. Molecular inflammation hypothesis of aging based on the anti-aging mechanism of calorie restriction. Microsc Res Tech. 2002 Nov 15;59(4):264-72. 22. Chung HY, Kim HJ, Kim JW, Yu BP. The inflammation hypothesis of aging: molecular modulation by calorie restriction. Ann N Y Acad Sci. 2001 Apr;928:327-35. 23. Kim DH, Kim JY, Yu BP, Chung HY. The activation of NF-kappaB through Akt-induced FOXO1 phosphorylation during aging and its modulation by calorie restriction. Biogerontology. 2008 Feb;9(1):33-47. 24. Yang SR, Wright J, Bauter M, Seweryniak K, Kode A, Rahman I. Sirtuin regulates cigarette smoke-induced proinflammatory mediator release via RelA/p65 NF-kappaB in macrophages in vitro and in rat lungs in vivo: implications for chronic inflammation and aging. Am J Physiol Lung Cell Mol Physiol. 2007 Feb;292(2):L567-76. 25. Pallas M, Casadesus G, Smith MA, et al. Resveratrol and neurodegenerative diseases: activation of SIRT1 as the potential pathway towards neuroprotection. Curr Neurovasc Res. 2009 Feb;6(1):70-81. 26. Kang OH, Jang HJ, Chae HS, et al. Anti-inflammatory mechanisms of resveratrol in activated HMC-1 cells: pivotal roles of NF-kappaB and MAPK. Pharmacol Res. 2009 May;59(5):330-7. 27. Park HJ, Jeong SK, Kim SR, et al. Resveratrol inhibits Porphyromonas gingivalis lipopolysaccharide-induced endothelial adhesion molecule expression by suppressing NF-kappaB activation. Arch Pharm Res. 2009 Apr;32(4):583-91. 28. Park JS, Kim KM, Kim MH, et al. Resveratrol inhibits tumor cell adhesion to endothelial cells by blocking ICAM-1 expression. Anticancer Res. 2009 Jan;29(1):355-62. 29. Roy P, Kalra N, Nigam N, et al. Resveratrol enhances ultraviolet B-induced cell death through nuclear factor-kappaB pathway in human epidermoid carcinoma A431 cells. Biochem Biophys Res Commun. 2009 Jun 26;384(2):215-20. 30. Rimando AM, Kalt W, Magee JB, Dewey J, Ballington JR. Resveratrol, pterostilbene, and piceatannol in vaccinium berries. J Agric Food Chem. 2004 Jul 28;52(15):4713-9. 31. Cichocki M, Paluszczak J, Szaefer H, Piechowiak A, Rimando AM, Baer-Dubowska W. Pterostilbene is equally potent as resveratrol in inhibiting 12-O-tetradecanoylphorbol-13-acetate activated NFkappaB, AP-1, COX-2, and iNOS in mouse epidermis. Mol Nutr Food Res. 2008 Jun;52 Suppl 1:S62-70. 32. Pan MH, Chiou YS, Chen WJ, Wang JM, Badmaev V, Ho CT. Pterostilbene inhibited tumor invasion via suppressing multiple signal transduction pathways in human hepatocellular carcinoma cells. Carcinogenesis. 2009 Jul;30(7):1234-42. 33. Priego S, Feddi F, Ferrer P, et al. Natural polyphenols facilitate elimination of HT-29 colorectal cancer xenografts by chemoradiotherapy: a Bcl-2- and superoxide dismutase 2-dependent mechanism. Mol Cancer Ther. 2008 Oct;7(10):3330-42. 34. Ruiz PA, Braune A, Holzlwimmer G, Quintanilla-Fend L, Haller D. Quercetin inhibits TNF-induced NF-kappaB transcription factor recruitment to proinflammatory gene promoters in murine intestinal epithelial cells. J Nutr. 2007 May;137(5):1208-15. 35. Min YD, Choi CH, Bark H, et al. Quercetin inhibits expression of inflammatory cytokines through attenuation of NF-kappaB and p38 MAPK in HMC-1 human mast cell line. Inflamm Res. 2007 May;56(5):210-5. 36. Terra X, Valls J, Vitrac X, et al. Grape-seed procyanidins act as antiinflammatory agents in endotoxin-stimulated RAW 264.7 macrophages by inhibiting NFkB signaling pathway. J Agric Food Chem. 2007 May 30;55(11):4357-65. 37. Chacon MR, Ceperuelo-Mallafre V, Maymo-Masip E, et al. Grape-seed procyanidins modulate inflammation on human differentiated adipocytes in vitro. Cytokine. 2009 Aug;47(2):137-42. 38. Ukil A, Maity S, Das PK. Protection from experimental colitis by theaflavin-3,3’-digallate correlates with inhibition of IKK and NF-kappaB activation. Br J Pharmacol. 2006 Sep;149(1):121-31. 39. Roy P, Nigam N, Singh M, et al. Tea polyphenols inhibit cyclooxygenase-2 expression and block activation of nuclear factor-kappa B and Akt in diethylnitrosoamine induced lung tumors in Swiss mice. Invest New Drugs. 2009 Jun 11. 40. Patel R, Krishnan R, Ramchandani A, Maru G. Polymeric black tea polyphenols inhibit mouse skin chemical carcinogenesis by decreasing cell proliferation. Cell Prolif. 2008 Jun;41(3):532-53. 41. Levites Y, Youdim MB, Maor G, Mandel S. Attenuation of 6-hydroxydopamine (6-OHDA)-induced nuclear factor-kappaB (NF-kappaB) activation and cell death by tea extracts in neuronal cultures. Biochem Pharmacol. 2002 Jan 1;63(1):21-9. 42. Cai F, Li C, Wu J, et al. Modulation of the oxidative stress and nuclear factor kappaB activation by theaflavin 3,3’-gallate in the rats exposed to cerebral ischemia-reperfusion. Folia Biol (Praha). 2007;53(5):164-72. 43. Chung HY, Cesari M, Anton S, et al. Molecular inflammation: underpinnings of aging and age-related diseases. Ageing Res Rev. 2009 Jan;8(1):18-30. 44. Kim YJ, Kim HJ, No JK, Chung HY, Fernandes G. Anti-inflammatory action of dietary fish oil and calorie restriction. Life Sci. 2006 Apr 18;78(21):2523-32. 45. Miliaras S, Anogeianaki A, Meditskou S, et al. Effects of rich-in-fat diets and highly selective COX-2 inhibitors on 7,12-dimethylbenz-(A)-anthracene-induced tumor growth. Int J Immunopathol Pharmacol. 2009 Apr-Jun;22(2):323-32. 46. Jung KJ, Lee EK, Kim JY, et al. Effect of short term calorie restriction on pro-inflammatory NF-kB and AP-1 in aged rat kidney. Inflamm Res. 2009 Mar;58(3):143-50. 47. Culpitt SV, Rogers DF, Fenwick PS, et al. Inhibition by red wine extract, resveratrol, of cytokine release by alveolar macrophages in COPD. Thorax. 2003 Nov;58(11):942-6. 48. Wu CT, Yu HP, Chung CY, Lau YT, Liao SK. Attenuation of lung inflammation and pro-inflammatory cytokine production by resveratrol following trauma-hemorrhage. Chin J Physiol. 2008 Dec 31;51(6):363-8. 49. Pan MH, Chang YH, Tsai ML, et al. Pterostilbene suppressed lipopolysaccharide-induced up-expression of iNOS and COX-2 in murine macrophages. J Agric Food Chem. 2008 Aug 27;56(16):7502-9. 50. de Souza Pereira R. Selective cyclooxygenase-2 (COX-2) inhibitors used for preventing or regressing cancer. Recent Pat Anticancer Drug Discov. 2009 Jun;4(2):157-63. 51. Tunon MJ, García-Mediavilla MV, Sánchez-Campos S, González-Gallego J. Potential of flavonoids as anti-inflammatory agents: modulation of pro-inflammatory gene expression and signal transduction pathways. Curr Drug Metab. 2009 Mar;10(3):256-71. 52. Cruz-Correa M, Shoskes DA, Sanchez P, et al. Combination treatment with curcumin and quercetin of adenomas in familial adenomatous polyposis. Clin Gastroenterol Hepatol. 2006 Aug;4(8):1035-8. 53. García-Mediavilla V, Crespo I, Collado PS,et al. The anti-inflammatory flavones quercetin and kaempferol cause inhibition of inducible nitric oxide synthase, cyclooxygenase-2 and reactive C-protein, and down-regulation of the nuclear factor kappaB pathway in Chang Liver cells. Eur J Pharmacol. 2007 Feb 28;557(2-3):221-9. 54. Sehirli O, Ozel Y, Dulundu E, Topaloglu U, Ercan F, Sener G. Grape seed extract treatment reduces hepatic ischemia-reperfusion injury in rats. Phytother Res. 2008 Jan;22(1):43-8. 55. Cai F, Li CR, Wu JL, et al. Theaflavin ameliorates cerebral ischemia-reperfusion injury in rats through its anti-inflammatory effect and modulation of STAT-1. Mediators Inflamm. 2006;2006(5):30490. 56. Alcain FJ, Villalba JM. Sirtuin inhibitors. Expert Opin Ther Pat. 2009 Mar;19(3):283-94. 57. Alcain FJ, Villalba JM. Sirtuin activators. Expert Opin Ther Pat. 2009 Apr;19(4):403-14. 58. Kanfi Y, Peshti V, Gozlan YM, Rathaus M, Gil R, Cohen HY. Regulation of SIRT1 protein levels by nutrient availability. FEBS Lett. 2008 Jul 9;582(16):2417-23. 59. Al-Regaiey KA, Masternak MM, Bonkowski MS, Panici JA, Kopchick JJ, Bartke A. Effects of caloric restriction and growth hormone resistance on insulin-related intermediates in the skeletal muscle. J Gerontol A Biol Sci Med Sci. 2007 Jan;62(1):18-26. 60. Fay JR, Steele V, Crowell JA. Energy homeostasis and cancer prevention: the AMP-activated protein kinase. Cancer Prev Res (Phila Pa). 2009 Apr;2(4):301-9. 61. Potente M, Dimmeler S. Emerging roles of SIRT1 in vascular endothelial homeostasis. Cell Cycle. 2008 Jul 15;7(14):2117-22. 62. Kritchevsky D. Caloric restriction and cancer. J Nutr Sci Vitaminol (Tokyo). 2001 Feb;47(1):13-9. 63. Pan MH, Chang YH, Tsai ML, et al. Pterostilbene suppressed lipopolysaccharide-induced up-expression of iNOS and COX-2 in murine macrophages. J Agric Food Chem. 2008 Aug 27;56(16):7502-9. 64. Ferrer P, Asensi M, Priego S, et al. Nitric oxide mediates natural polyphenol-induced Bcl-2 down-regulation and activation of cell death in metastatic B16 melanoma. J Biol Chem. 2007 Feb 2;282(5):2880-90. 65. Pan MH, Chang YH, Badmaev V, Nagabhushanam K, Ho CT. Pterostilbene induces apoptosis and cell cycle arrest in human gastric carcinoma cells. J Agric Food Chem. 2007 Sep 19;55(19):7777-85. 66. Pan MH, Lin YT, Lin CL, Wei CS, Ho CT, Chen WJ. Suppression of Heregulin-beta1/HER2-Modulated Invasive and Aggressive Phenotype of Breast Carcinoma by Pterostilbene via Inhibition of Matrix Metalloproteinase-9, p38 Kinase Cascade and Akt Activation. Evid Based Complement Alternat Med. 2009 Jul 16. 67. Hsieh TC. Uptake of resveratrol and role of resveratrol-targeting protein, quinone reductase 2, in normally cultured human prostate cells. Asian J Androl. 2009 Sep 21. 68. Gescher AJ. Resveratrol from red grapes - pedestrian polyphenol or useful anticancer agent? Planta Med. 2008 Oct;74(13):1651-5. 69. Chien SY, Wu YC, Chung JG, et al. Quercetin-induced apoptosis acts through mitochondrial- and caspase-3-dependent pathways in human breast cancer MDA-MB-231 cells. Hum Exp Toxicol. 2009 Aug;28(8):493-503. 70. Kang JW, Kim JH, Song K, Kim SH, Yoon JH, Kim KS. Kaempferol and quercetin, components of Ginkgo biloba extract (EGb 761), induce caspase-3-dependent apoptosis in oral cavity cancer cells. Phytother Res. 2009 Jul 7. 71. Tan J, Wang B, Zhu L. Regulation of survivin and Bcl-2 in HepG2 cell apoptosis induced by quercetin. Chem Biodivers. 2009 Jul;6(7):1101-10. 72. Kaindl U, Eyberg I, Rohr-Udilova N, Heinzle C, Marian B. The dietary antioxidants resveratrol and quercetin protect cells from exogenous pro-oxidative damage. Food Chem Toxicol. 2008 Apr;46(4):1320-6. 73. Kaur M, Singh RP, Gu M, Agarwal R, Agarwal C. Grape seed extract inhibits in vitro and in vivo growth of human colorectal carcinoma cells. Clin Cancer Res. 2006 Oct 15;12(20 Pt 1):6194-202. 74. Wen W, Lu J, Zhang K, Chen S. Grape seed extract inhibits angiogenesis via suppression of the vascular endothelial growth factor receptor signaling pathway. Cancer Prev Res (Phila Pa). 2008 Dec;1(7):554-61. 75. Letchoumy PV, Mohan KV, Prathiba D, Hara Y, Nagini S. Comparative evaluation of antiproliferative, antiangiogenic and apoptosis inducing potential of black tea polyphenols in the hamster buccal pouch carcinogenesis model. J Carcinog. 2007;6:19. 76. Prasad S, Kaur J, Roy P, Kalra N, Shukla Y. Theaflavins induce G2/M arrest by modulating expression of p21waf1/cip1, cdc25C and cyclin B in human prostate carcinoma PC-3 cells. Life Sci. 2007 Oct 13;81(17-18):1323-31. 77. Ugochukwu NH, Figgers CL. Modulation of the flux patterns in carbohydrate metabolism in the livers of streptozoticin-induced diabetic rats by dietary caloric restriction. Pharmacol Res. 2006 Sep;54(3):172-80. 78. Liang F, Kume S, Koya D. SIRT1 and insulin resistance. Nat Rev Endocrinol. 2009 Jul;5(7):367-73. 79. Carvajal K, Hernández-Esquivel Mde L, Moreno-Sánchez R. PPARs, metabolic syndrome and cardiac diseases. Arch Cardiol Mex. 2007 Oct-Dec;77 Suppl 4:S4-66-76. 80. Cheng G, Zhang X, Gao D, Jiang X, Dong W. Resveratrol inhibits MMP-9 expression by up-regulating PPAR alpha expression in an oxygen glucose deprivation-exposed neuron model. Neurosci Lett. 2009 Feb 20;451(2):105-8. 81. Mizuno CS, Ma G, Khan S, Patny A, Avery MA, Rimando AM. Design, synthesis, biological evaluation and docking studies of pterostilbene analogs inside PPARalpha. Bioorg Med Chem. 2008 Apr 1;16(7):3800-8. 82. Rimando AM, Nagmani R, Feller DR, Yokoyama W. Pterostilbene, a new agonist for the peroxisome proliferator-activated receptor alpha-isoform, lowers plasma lipoproteins and cholesterol in hypercholesterolemic hamsters. J Agric Food Chem. 2005 May 4;53(9):3403-7. 83. Floyd ZE, Wang ZQ, Kilroy G, Cefalu WT. Modulation of peroxisome proliferator-activated receptor gamma stability and transcriptional activity in adipocytes by resveratrol. Metabolism. 2008 Jul ;57(7 Suppl 1):S32-8. 84. Kennedy A, Overman A, Lapoint K, et al. Conjugated linoleic acid-mediated inflammation and insulin resistance in human adipocytes are attenuated by resveratrol. J Lipid Res. 2009 Feb;50(2):225-32. 85. Lopez-Lluch G, Irusta PM, Navas P, de Cabo R. Mitochondrial biogenesis and healthy aging. Exp Gerontol. 2008 Sep;43(9):813-9 86. Pinent M, Blade MC, Salvado MJ, Arola L, Ardevol A. Intracellular mediators of procyanidin-induced lipolysis in 3T3-L1 adipocytes. J Agric Food Chem. 2005 Jan 26;53(2):262-6. 87. Pinent M, Blade MC, Salvado MJ, et al. Grape-seed derived procyanidins interfere with adipogenesis of 3T3-L1 cells at the onset of differentiation. Int J Obes (Lond). 2005 Aug;29(8):934-41. 88. Ma L, Gao HQ, Li BY, Ma YB, You BA, Zhang FL. Grape seed proanthocyanidin extracts inhibit vascular cell adhesion molecule expression induced by advanced glycation end products through activation of peroxisome proliferators-activated receptor gamma. J Cardiovasc Pharmacol. 2007 May;49(5):293-8. 89. Liu Y, Dentin R, Chen D, et al. A fasting inducible switch modulates gluconeogenesis via activator/coactivator exchange. Nature. 2008 Nov 13;456(7219):269-73. 90 Palsamy P, Subramanian S. Modulatory effects of resveratrol on attenuating the key enzymes activities of carbohydrate metabolism in streptozotocin-nicotinamide-induced diabetic rats. Chem Biol Interact. 2009 May 15;179(2-3):356-62. 91. Pari L, Satheesh MA. Effect of pterostilbene on hepatic key enzymes of glucose metabolism in streptozotocin- and nicotinamide-induced diabetic rats. Life Sci. 2006 Jul 10;79(7):641-5. 92. Pinent M, Blay M, Blade MC, Salvado MJ, Arola L, Ardevol A. Grape seed-derived procyanidins have an antihyperglycemic effect in streptozotocin-induced diabetic rats and insulinomimetic activity in insulin-sensitive cell lines. Endocrinology. 2004 Nov;145(11):4985-90. 93. Vessal M, Hemmati M, Vasei M. Antidiabetic effects of quercetin in streptozocin-induced diabetic rats. Comp Biochem Physiol C Toxicol Pharmacol. 2003 Jul;135C(3):357-64. 94. Ramana BV, Raju TN, Kumar VV, Reddy PU. Defensive role of quercetin against imbalances of calcium, sodium, and potassium in galactosemic cataract. Biol Trace Elem Res. 2007 Oct;119(1):35-41. 95. Kusano R, Andou H, Fujieda M, Tanaka T, Matsuo Y, Kouno I. Polymer-like polyphenols of black tea and their lipase and amylase inhibitory activities. Chem Pharm Bull (Tokyo). 2008 Mar;56(3):266-72. 96. Kobayashi M, Ichitani M, Suzuki Y,et al. Black-tea polyphenols suppress postprandial hypertriacylglycerolemia by suppressing lymphatic transport of dietary fat in rats. J Agric Food Chem. 2009 Aug 12;57(15):7131-6. 97. Shoji Y, Nakashima H. Glucose-lowering effect of powder formulation of African black tea extract in KK-A(y)/TaJcl diabetic mouse. Arch Pharm Res. 2006 Sep;29(9):786-94. 98. Ramadan G, El-Beih NM, Abd El-Ghffar EA. Modulatory effects of black v. green tea aqueous extract on hyperglycaemia, hyperlipidaemia and liver dysfunction in diabetic and obese rat models. Br J Nutr. 2009 Oct 13:1-9. 99. Cameron AR, Anton S, Melville L, et al. Black tea polyphenols mimic insulin/insulin-like growth factor-1 signalling to the longevity factor FOXO1a. Aging Cell. 2008 Jan;7(1):69-77. 100. Morris BJ. A forkhead in the road to longevity: the molecular basis of lifespan becomes clearer. J Hypertens. 2005 Jul;23(7):1285-3. |

Wellness
Specialists
1-800-226-2370 - This service is FREE
7:30 AM - 12 AM (ET) Mon-Fri | 9 AM - 12 AM (ET) Sat-Sun