Life Extension Magazine®

How Immune Decline Hastens Aging
Few people realize that immune degeneration predisposes us to cancer, vascular disease, infection, and senility, along with inflammatory disorders that shorten our life span. Researchers have identified specific immune components that decline with aging and have discovered natural ways to restore more youthful immune profiles in maturing individuals.
Scientifically reviewed by Dr. Gary Gonzalez, MD, in October 2024. Written by: William Faloon.

This has been an incredible year of scientific achievement. Even the lay public is coming to the realization that profoundly extended health spans may be just around the corner.
A concern expressed by our older members is whether they can remain alive long enough to benefit from the upcoming biomedical revolution. These health activists want to determine their areas of vulnerability to ensure they’re not overlooking a validated way to stave off disease, aging, and death.
Our work involving longevity research dates back to the 1960s. This has enabled us to gain exclusive insights into what causes people to die—people who otherwise take exceptionally good care of themselves.
A sad example of mortality vulnerability occurred in January 2014. That’s when one of our pioneer members died at the young age of 92. His intellectual contributions, spanning many decades, helped us survive withering governmental assaults aimed at censoring our ability to disseminate lifesaving information.
I can’t stop thinking that we were only weeks away from potentially saving him. I don’t want this same fate to befall other Life Extension® members.
We’ve talked before about the lethal impact of immune senescence. It occurs when our aging immune system fails to protect against cancers/infections, and instead generates excess inflammatory reactions that attack every cell in our body.
What virtually no one understands is how aging accelerates immune decline and what must be done to reverse this lethal trend.
The most common afflictions associated with normal aging are atherosclerosis, cancer, and Alzheimer’s disease. 1 What few doctors know is that all these illnesses are related to immune senescence.2,3
In people over age 65, the top 10 causes of death include pneumonia, influenza, and sepsis (systemic inflammation caused by severe infection).1 Immune senescence is a major cause of all these maladies.4-10
As winter sets in, the term “immune system” is liberally tossed around as people seek to protect against viral infections. What the public does not yet understand is what causes our immune system to fail as we age.11
Why Young Children Catch So Many Colds
Young children get lots of colds. Some suffer as many as eight to ten each year before age 2.12
The reason youngsters suffer more colds than older children and adults is because they haven’t built up “immunity” to cold viruses.12 But what does this immunity really mean?
When one is exposed to an infectious agent, the body creates and maintains “memory T-cells” that provide a degree of immunity to the same infection.13 This works magnificently well in our first four to five decades of life. But then something happens that leads to the downward spiral of immune senescence.
Too Many “Memory T-Cells”
Memory T-cells form when our immune system successfully eradicates an invader. They remain in the body and are ready to instantly respond when that same bacteria, virus, or cancer cell reappears.
One would think it desirable to have lots of memory T-cells. The problem is that memory T-cells only work against prior infections. As we age, we collect excess numbers of memory T-cells and produce fewer critically important naïve T-cells.14
The term “naïve” may not sound like something beneficial as it relates to immune function, but it is. A naïve immune cell is one that has not been activated by an antigen (a substance that provokes an adaptive immune response). Since it is “ naïve” (not yet exposed to an antigen), naïve immune cells are primed to effectively respond to new infectious agents and malignancies.15
Once exposed, naïve immune cells become memory cells or plasma cells specific to the original antigen. As our internal reservoir of naïve immune cells is decreased, we have less ability to respond to new infections and malignancies.16
To make matters worse, excess numbers of senescent memory cells provoke undesirable inflammatory reactions 17-21 that are thought to underlie most age-related diseases including atherosclerosis, cancer, and dementia.22-26
To put this in simplistic terms, if we are to protect against the ravages of immune senescence, we need to increase our numbers of naïve cells (“virgin” immune cells), while reducing numbers of surplus senile memory cells.
Importance Of “Functional” Natural Killer Cells
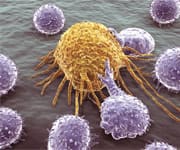
The first line of defense against virus-infected28-31 and cancer cells is our natural killer cells.32-45 Young individuals have high levels of functional natural killer (NK) immune cells, but this declines with aging. 46-49
In elderly subjects, decreased NK cell activity is associated with an increased incidence and severity of viral infections such as shingles, influenza, and cytomegalovirus (CMV). 47,50-53
Shingles occurs when our immunity to dormant chickenpox viral infection declines. It manifests as an extremely painful skin lesion that can last for months. 54
Influenza, commonly called the flu, is a virus that inflicts its lethal effects mostly on the elderly, who represent the largest portion of the population that suffers immune dysfunction.1,7
Cytomegalovirus (CMV) is a chronic infection that, as you’ll discover later in this issue, may contribute significantly to degenerative disease. About 90% of older people showed history of CMV infection on lab testing compared to about 60% of the general population. 55 Increased prevalence of CMV in the elderly is thought to lead to decreased immune surveillance.56-61
Natural killer cells originate in the bone marrow (like other immune cells) and go through a maturation process that enables them to participate in early control of microbial infections and cancers.47,62
Healthy NK function is critical in eliminating transformed cancer cells.27,44,63 NK cells are also involved in the elimination of senescent cells62,64 that otherwise cause chronic inflammation.27,65-67
The age-related decrease in functional NK cells is likely to have wider implications for the health of older adults than currently understood by the mainstream. If an aging person is to better manage debilitating and deadly infections and malignancies, maintaining youthful NK function is critical.
 |
© Nucleus Medical Art, Inc / Phototake
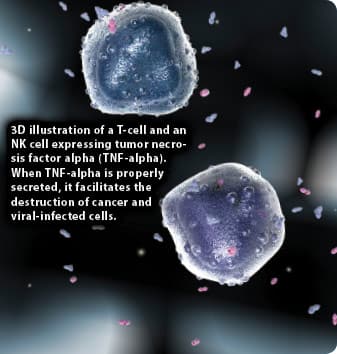
“In elderly subjects, decreased NK cell activity has been shown to be associated with an increased incidence and severity of viral infection, highlighting the clinical implications that age-associated changes in NK cell biology have on the health of older adults… Evidence has emerged that has shown that in addition to eliminating transformed cells, NK cells are involved in many other biological processes such as immune regulation, antimicrobial immune responses, and the recognition and elimination of senescent cells… Thus, the decrease in NK cell function that accompanies physiological aging is likely to have wider implications for the health of older adults than originally thought.” 27
Reference: Aging Research Reviews-May 6, 2013. “The Impact of Aging on Natural Killer Cell Function and Potential Consequences for Health in Older Adults.”
Consequences Of Immune Cell Exhaustion
Over the course of our lifetime, our immune system becomes “exhausted.” What this refers to is the excess accumulation of worn-out memory T-cells and reduced production of vital naïve T-cells.68-70
As people accumulate exhausted T-cells, an adverse consequence is these senile memory cells emit pro-inflammatory cytokines22,71 that exacerbate chronic inflammatory conditions.72,73 Individuals with higher levels of exhausted immune cells suffer greater mortality.74,75
The deficit of naïve immune cells combined with over accumulation of exhausted memory cells decreases the efficacy (antibody response) of vaccinations.76-78
Exhausted memory T-cells are associated with increased inflammation.79 Inflammation is associated with increased risk of coronary heart disease, impaired vascular function, vascular inflammation, and endothelial dysfunction.80-83
An accumulation of exhausted T-cells has been seen in persons suffering from rheumatoid arthritis84 and ankylosing spondylitis (an inflammatory disease of the spine).85-87
Compelling evidence points to the accumulation of senile (exhausted) T-cells as a factor that accelerates a broad array of age-associated diseases.71
 |
“We conclude that chronic heart failure patients show a higher degree of immunosenescence than age-matched healthy controls. T-lymphocyte differentiation and interleukin-6 (IL-6) levels are increased in patients with an advanced clinical status and may contribute to disease impairment through a compromised adaptive immune response due to accelerated aging of their immune system.” 88
Reference: International Journal of Cardiology-July 1, 2014. “Immunosenescence and Inflammation Characterize Chronic Heart Failure Patients With More Advanced Disease.”
More T-Helper Cells Needed
T-helper cells identify and tag invaders for elimination by the immune system.89
Regulatory T-cells tell the immune system that its job is finished and it’s time to stop the attack.89-91 A normal balance involves having at least one to four T-helper cells for each regulatory T- cell (regulatory T-cells are sometimes called suppressor T-cells).92
As humans age, there is an inversion in the T-helper/regulatory T-cell ratio.93 What this means is that too many regulatory T-cells form while T-helper cell counts drop, resulting in there being more regulatory T-cells than T-helpers. The T-helper/regulatory T-cell ratio can be considered to be a predictor of mortality.76,77,94 People with low T-helper counts and higher regulatory T-cell counts die sooner.
Cancer patients often present with a low T-helper/regulatory T-cell ratio.95,96 Some studies show that tumor cells secrete chemicals that turn up regulatory T-cell formation in order to prevent the immune system from attacking cancer cells.97-99 Cancer chemotherapy dramatically lowers T-helper counts.100,101
To combat immune senescence, it is critical to reverse the inversion of the T-helper/regulatory T-cell ratio. This means boosting T-helper counts while lowering regulatory T-cells.
Hidden Effects Of Natural Killer (NK) Cell Senescence
 |
Natural killer (NK) cells are best known for their impact on reducing the incidence and severity of viral infections.28-31
Overlooked are studies showing vibrant NK function is crucial for the immune system to recognize and eliminate unhealthy/infected cells and dysfunctional (senile) cells.27,115 A pathologic consequence of too many senile cells is tissue/organ damage and chronic inflammation.116-119
Senile (senescent) cells reside in a state of irreversible cellular paralysis. Your body wants them to die and disappear, but they linger on for dangerously long periods. While these senile cells contribute nothing to healthy function, they remain metabolically active, secreting an array of dangerous growth factors and pro-inflammatory cytokines.
Functional NK cells can eliminate senile cells before they buildup to lethal levels. When NK cells are hampered by aging, however, they are less able to remove deadly senile cells from our bodies.
As NK cells themselves become senescent in older people, there is increased reactivation of tuberculosis and viral illnesses (such as shingles and CMV), slow resolution of inflammatory responses, and increased incidence of bacterial/fungal infections and malignancies. 27,120
It is likely that the senescence of NK cells has wider implications on the health of older adults than originally thought. NK cell senescence thus represents a dagger of vulnerability that predisposes us to premature suffering and death. It must be reversed to avert degenerative disease.
Reversing Immune Senescence
Immune senescence is a prime cause of debility and mortality.
Fortunately, discoveries in recent years have been shown to help reverse the immune dysfunction that plagues virtually all aging humans (and cancer patients).
Here is a summary of the six underlying causes of immune senescence described in this article:
- Decrease in naïve immune T-cells needed to fight new invaders.93
- Increase in exhausted memory T-cells that create chronic inflammatory reactions.93
- Decrease in functional natural killer (NK) cell activity.93
- Thymus gland atrophy that reduces T-cell function and numbers.
- Too many regulatory T-cells and a reduction in T-helper cells.
- Excess production of interleukin-6, a cytokine that promotes inflammation.102
The good news is that there are proven ways to counteract all six factors involved in senile immune dysfunction.
Most Life Extension members already take zinc and DHEA. There is evidence that these supplements can at least partially restore thymic function vital to transforming immune cells produced in bone marrow to mature T-cells.103-105 DHEA also helps suppress deadly interleukin-6.106,107
A breakthrough in combatting immune senescence has been found in a medicinal plant called cistanche that has been used extensively in China to treat the “ailments of aging.”108 Until recently, doctors would not have understood how cistanche reverses some of the underlying causes of immune senescence.
Supplementation with cistanche has been shown to increase naïve T-cells and natural killer (NK) cells while decreasing memory T-cells and pro-inflammatory interleukin-6.109
One of the characteristics of people who live over 100 years is a low level of interleukin-6.
A prime reason for the severe immune disorders suffered by the elderly is the marked decrease in naïve T-cells and functional natural killer cells, with a concomitant increase in memory T-cells. Cistanche reverses these pathological trends that characterize immune senescence.
One way cistanche functions is by restoring the progenitors of peripheral naïve T-cells, which explains the increase seen in these vital cells in response to cistanche.109 Animals supplemented with cistanche have increased life spans, as would be expected by a compound that counteracts immune senescence.109
Cistanche is one of the most popular Chinese herbal medicines and is listed in the Chinese herbal pharmacopeias as having “anti-aging” properties.
One reason the Chinese saw such impressive therapeutic results is that cistanche restores one of the most prominent bone marrow biomarkers of immune cell formation called stem cell antigen-1.109 Senile bone marrow loses its ability to produce fresh naïve immune cells, which are launched into the bloodstream to differentiate into mature naïve T and natural killer cells. Bone marrow stem cell antigen-1 represents the body’s main source of naïve T-cells in the blood.109 Cistanche appears to have a rejuvenating effect on the bone marrow, something that is now only possible using very expensive recombinant drugs.110-112
There are other factors that weaken immune function in the elderly that cistanche has been shown to counteract. These will be described in depth in an article appearing in this issue.
Perhaps the most exciting finding was an open-label pilot trial of elderly people that combined a low-dose of cistanche (100 mg) with zinc, vitamin E, vitamin B6, fucoidan, and coenzyme Q10. Not only were markers of immune senescence reversed, but the test subjects reported improvements in quality of life, such as not “feeling tired all the time.” This makes sense in light of the multiple adverse effects immune senescence inflicts on the body, which includes increased levels of frailty. 113,114
Most People Don’t Get Enough Zinc
 |
Immune cells are produced in the bone marrow and programmed into functional T-cells by our thymus gland. Profound shrinkage (atrophy) of the thymus gland occurs with aging and in the presence of nutrient deficiency—especially zinc.121,122
Zinc supplementation in old animals stimulates thymus gland growth and increases youthful thymic functions critical to healthy immune regulation. 123,124
In older adult humans, zinc deficiency is rampant. One reason for this is that the government’s minimum recommended daily allowance for zinc is only 8 to 11 mg.125 Yet 35 to 45% of people over age 60 don’t even get half of that.126,127
Optimal levels of supplemental zinc is around 30 to 50 mg a day, so it is easy to see why even people taking basic RDA-potency vitamins are woefully short of this critical mineral.
Scientists now believe that zinc deficiency plays a direct role in the aging of the immune system.128,129 The hormone DHEA is also important in maintaining youthful thymic function.
Zinc and DHEA are remarkably low-cost dietary supplements, and the fact that most people are deficient in one or both of them helps explain the epidemic of degenerative disease that occurs as human’s age past 60 years, which is when immune senescence significantly manifests. As you’ve been reading, however, more than just zinc and DHEA are needed to ward off immune senescence.
How To Take Advantage Of Novel Immune Restorative Technologies
Unlike prescription drugs or even certain dietary supplements, cistanche does not cost a lot. It has been added to the most popular nutrient members now take to bolster immune function (Reishi). Virtually every person over 35 should add this cistanche/Reishi immune protection duo to their daily regimen.
Another supplement to consider for short-term use is enzymatically modified rice bran, which has been demonstrated to have overwhelming benefits in boosting natural killer cell activity.130-134 The problem we have with this supplement is its high cost. What we are going to suggest, and will provide a rationale for, is for members to take this enzymatically modified rice bran for only four months out of the year.134
We believe the substantial boost in natural killer cell activity will help eliminate virus-infected cells, premalignant cells, and senile cells that linger in the body and emit constant streams of pro-inflammatory cytokines. An underlying cause of aging is the accumulation of senile (senescent) cells that fail to undergo apoptosis (natural cell cycle destruction).135 An increase in natural killer (NK) activity can help purge the body of these unwanted senescent cells. You’re also going to learn in this issue of Life Extension magazine why removing cells infected with the CMV (cytomegalovirus) can result in a meaningful extension of the healthy human life span.
To further restore a more youthful immune profile, we’re recommending that members go on a 60-day course of an over-the-counter drug called cimetidine.
This can be purchased at pharmacies at very low cost. We suggest that most people take 800 mg of cimetidine each night for 60 consecutive days to reduce excess regulatory T-cell counts and increase T-helper cell counts. Regulatory T-cells sometimes turn off the immune system before immune eradication of virus-infected cells and tumor cells occurs.
We think this 60-day cycle of cimetidine should be considered a few times during the year, but do not take it all the time. We don’t have the data yet to support everyday use of cimetidine in protecting against immune senescence, but there is compelling evidence for shorter-term use.136 Cimetidine is approved by the FDA for use in heartburn sufferers (The brand name of this drug is Tagamet®.)
A trial was done on colorectal cancer patients who used 800 mg a day of cimetidine for one year. In these patients, cimetidine demonstrated significant survival benefits—84.6% of the cimetidine/fluorouracil group were alive after 10 years compared to only 49.8% of the control group (given fluorouracil alone).137
Life Extension has recommended cimetidine to certain cancer patients since 1985, and the most robust benefits have been shown to occur when cimetidine is administered prior to surgery and other immunosuppressive cancer therapies.138-142
We urge members to get on a 60-day cycle of cimetidine now to bolster defenses against immune senescence and winter infections.
In another article of this issue is a listing of cimetidine side effects and contraindications.
We’re On The Verge Of Something Big…
In case you have not figured it out yet, we are entering a new paradigm in the prevention and reversal of age-related disease.
While certain nutrients Life Extension members have taken for decades help protect against immune decline, never before have we had such an arsenal to counteract the multiple underlying factors that characterize immune senescence. And this is just the beginning.
We are funding aggressive clinical research involving bone marrow rejuvenation with the objective of mobilizing hematopoietic stem and progenitor cells (HSPCs) that will rejuvenate every tissue of our aging bodies.
We are going even further in helping the very elderly with a research project that involves taking stem cell-stimulated blood of individuals under age 25, tissue-type matching it to elderly individuals, separating the youth factors from this young blood, and transfusing it into the elderly subjects. The objective of this research project is nothing short of meaningful age reversal in individuals who may have only a few more months or years to live.
How is this expensive research funded? By the dietary supplements you purchase from the Life Extension Buyers Club. We use proceeds from supplement sales to fund a wide range of projects aimed at finding cures for cancer, vascular disease, aging, and death itself. A record number of media stories this year highlighted areas of research we pioneered that are now being studied at prestigious universities.
Premature Death Of Jack LaLanne Likely Caused By Immune Senescence
 |
It is impossible to overstate the magnitude of disability and death that immune senescence inflicts on aging human populations. When you hear a person died from “old age,” in almost every instance the underlying culprit is a dysfunctional immune system that ignites deadly inflammatory fires while failing to protect against infections and malignancies.
No one we know tried to live for over 100 years more than health pioneer Jack LaLanne. He engaged in rigorous lifelong exercise and ate a low-calorie healthy diet. He followed an old-line supplement program probably devoid of vitamin K and DHEA, which may have contributed to his premature death at age 96.
Jack LaLanne died of pneumonia following aortic valve surgery. Aortic valve stenosis is caused by calcification, chronic inflammation, and other factors such as elevated homocysteine.143
Vitamin K prevents calcification that is so often the reason why an elderly person’s aortic valve fails.144-149 DHEA partially protects against inflammation.150-152
Immune senescence may very well have contributed to Jack LaLanne’s early demise by crippling his ability to fight off the pneumonia, which was the acute cause of his death.
It is difficult to imagine someone as fit and vigorous as Jack LaLanne succumbing to pneumonia. Yet a dysfunctional immune system deprives even the hardiest of us from warding off infections that were easily overcome in our youth.
Clearly, protecting against immune senescence is a mandatory component of a longevity program.
Obtain Latest Formulations At Discount Prices
Once a year, we discount the price of every one of our advanced nutritional formulas. Members take advantage of this once-a-year Super Sale to stock up on their favorite supplement formulas.
What should comfort members more than anything else are the three novel methods to counteract aging we have introduced over the past few months. In addition to the immune senescence reversal program discussed in this editorial, there is now a supplement that mimics several of the longevity properties of the drug metformin, and a cell-regenerating nutrient called nicotinamide riboside that we’ve been working on since the year 2001.
We’ve also combined some nutrients into existing formulas so that members can obtain more benefits while swallowing fewer pills.
No organization in the world is combatting senescence and human mortality more aggressively than Life Extension. Your support via supplement purchases and donations enables us to expand on an unprecedented biomedical research endeavor.
To order nutrients you need today at Super Sale prices, call 1-800-544-4440.
For longer life,
William Faloon
References
- Available at: http://www.cdc.gov/nchs/data/ahcd/agingtrends/01death.pdf. Accessed October 24, 2014.
- Tsoukas P. Immune senescence and cardiovascular morbidity as a result of chronic cytomegalovirus infection. RCSIsmj. 2012;5:67-70.
- Fulop T, Fortin C, Lesur O, et al. The innate immune system and aging: What is the contribution to immunosenescence? Open Longevity Science. 2012;6:121-32.
- Shivshankar P, Boyd AR, Le Saux CJ, Yeh IT, Orihuela CJ. Cellular senescence increases expression of bacterial ligands in the lungs and is positively correlated with increased susceptibility to pneumococcal pneumonia. Aging Cell. 2011 Oct;10(5):798-806.
- Bartling B. Cellular senescence in normal and premature lung aging. Z Gerontol Geriatr. 2013 Oct;46(7):613-22.
- Glennie SJ, Sepako E, Mzinza D, et al. Impaired CD4 T-cell memory response to Streptococcus pneumoniae precedes CD4 T-cell depletion in HIV-infected Malawian adults. PLoS One. 2011 6(9):e25610.
- Hernandez-Vargas EA, Wilk E, Canini L, et al. Effects of aging on influenza virus infection dynamics. J Virol. 2014 Apr;88(8):4123-31.
- Zhou X, McElhaney JE. Age-related changes in memory and effector T-cells responding to influenza A/H3N2 and pandemic A/H1N1 strains in humans. Vaccine. 2011 Mar 3;29(11):2169-77.
- Opal SM, Girard TD, Ely EW. The immunopathogenesis of sepsis in elderly patients. Clin Infect Dis. 2005 Nov 15;41 Suppl 7:S504-12.
- Vollmar B, Pradarutti S, Nickels RM, Menger MD. Age-associated loss of immunomodulatory protection by granulocyte-colony stimulating factor in endotoxic rats. Shock. 2002 Oct;18(4):348-54.
- Mekker A, Tchang VS, Haeberli L, Oxenius A, Trkola A, Karrer U. Immune senescence: Relative contributions of age and cytomegalovirus infection. PLoS Pathog. 2012;8(8):e1002850.
- [No authors listed]. Colds in children. Paediatr Child Health. 2005 Oct;10(8):493-5.
- Verhoeven D, Teijaro JR, Farber DL. Heterogeneous memory T-cells in antiviral immunity and immunopathology. Viral Immunol. 2008 Jun;21(2):99-113.
- Berard M, Tough DF. Qualitative differences between naive and memory T-cells. Immunology. 2002 June;106(2):127-38.
- Whitmire JK, Eam B, Whitton JL. Tentative T-cells: memory cells are quick to respond, but slow to divide. PLoS Pathog. 2008 Apr 11;4(4):e1000041.
- Janeway CA Jr, Travers P, Walport M, et al. Immunobiology: The Immune System in Health and Disease. 5th edition. New York: Garland Science; 2001. Chapter 8; T-cell-Mediated Immunity.
- Norrie IC, Ohlsson E, Nielsen O, Hasemann MS, Porse BT. C/EBPα is dispensable for the ontogeny of PD-1+ CD4+ memory T-cells but restricts their expansion in an age-dependent manner. PLoS One. 2014 Jan 3;9(1):e84728.
- Pawelec G, Larbi A, Derhovanessian E. Senescence of the human immune system. J Comp Pathol. 2010 Jan;142 Suppl 1: S39-44.
- Vallejo AN, Weyand CM, Goronzy JJ. T-cell senescence: a culprit of immune abnormalities in chronic inflammation and persistent infection. Trends Mol Med. 2004 Mar;10(3):119-24.
- Franceschi C, Bonafè M, Valensin S, et al. Inflamm-aging. An evolutionary perspective on immunosenescence. Ann N Y Acad Sci. 2000 Jun;908:244-54.
- Freund A, Orjalo AV, Desprez PY, Campisi J. Inflammatory networks during cellular senescence: causes and consequences. Trends Mol Med. 2010 May;16(5):238-46.
- Chou JP, Effros RB. T-cell replicative senescence in human aging. Curr Pharm Des. 2013;19(9):1680-98.
- Sikora E, Bielak-Zmijewska A, Mosieniak G. Cellular Senescence in Ageing, Age-Related Disease and Longevity. Curr Vasc Pharmacol. 2013 Dec 18.
- Shaw AC, Goldstein DR, Montgomery RR. Age-dependent dysregulation of innate immunity. Nat Rev Immunol. 2013 Dec;13(12):875-87.
- Falci C, Gianesin K, Sergi G, et al. Immune senescence and cancer in elderly patients: results from an exploratory study. Exp Gerontol. 2013 Dec;48(12):1436-42.
- Giunta B, Fernandez F, Nikolic WV, et al. Inflammaging as a prodrome to Alzheimer’s disease. J Neuroinflammation. 2008 Nov 11;5:51.
- Hazeldine J, Lord JM. The impact of ageing on natural killer cell function and potential consequences for health in older adults. Ageing Res Rev. 2013 Sep;12(4):1069-78.
- Wang D, Ma Y, Wang J, Liu X, Fang M (2013) Natural killer cells in innate defense against infective pathogens. J Clin Cell Immunol. S13:006.
- Lodoen MB, Lanier LL. Natural killer cells as an initial defense against pathogens. Curr Opin Immunol. 2006 Aug;18(4):391-8.
- Brandstadter JD, Yang Y. Natural killer cell responses to viral infection. J Innate Immun. 2011;3(3):274-9.
- Orange JS. Human natural killer cell deficiencies and susceptibility to infection. Microbes Infect. 2002 Dec;4(15):1545-58.
- Arina A, Murillo O, Dubrot J, et al. Cellular liaisons of natural killer lymphocytes in immunology and immunotherapy of cancer . Expert Opin Biol Ther. 2007 May;7(5):599-615.
- Terunuma H, Deng X, Dewan Z, Fujimoto S, Yamamoto N. Potential role of NK cells in the induction of immune responses: implications for NK cell-based immunotherapy for cancers and viral infections. Int Rev Immunol. 2008;27(3):93-110.
- Wiltrout RH, Herberman RB, Zhang SR, et al. Role of organ-associated NK cells in decreased formation of experimental metastases in lung and liver. J Immunol. 1985 Jun;134(6):4267-75.
- Arai S, Meagher R, Swearingen M, et al. Infusion of the allogeneic cell line NK-92 in patients with advanced renal cell cancer or melanoma: a phase I trial. Cytotherapy. 2008;0(6):625-32.
- Tonn T, Becker S, Esser R, Schwabe D, Seifried E. Cellular immunotherapy of malignancies using the clonal natural killer cell line NK-92. J Hematother Stem Cell Res. 2001 Aug;10(4):535-44.
- Oberoi P, Wels WS. Arming NK cells with enhanced antitumor activity: CARs and beyond. Oncoimmunology. 2013 Aug 1;2(8):e25220.
- Ziske C, Märten A, Schöttker B, et al. Resistance of pancreatic carcinoma cells is reversed by co-culturing NK-like T-cells with dendritic cells pulsed with tumor-derived RNA and CA 19-9. Mol Ther. 2001 Jan;3(1):54-60.
- Villegas FR, Coca S, Villarrubia VG, et al. Prognostic significance of tumor infiltrating natural killer cells subset CD57 in patients with squamous cell lung cancer. Lung Cancer. 2002 Jan;35(1):23-8.
- Ishigami S, Natsugoe S, Tokuda K, et al. Prognostic value of intratumoral natural killer cells in gastric carcinoma. Cancer. 2000 Feb 1;88(3):577-83.
- Coca S, Perez-Piqueras J, Martinez D, et al. The prognostic significance of intratumoral natural killer cells in patients with colorectal carcinoma. Cancer. 1997 Jun 15;79(12):2320-8.
- Pietra G, Manzini C, Vitale M, et al. Natural killer cells kill human melanoma cells with characteristics of cancer stem cells. Int Immunol. 2009 Jul;21(7):793-801.
- Castriconi R, Daga A, Dondero A, et al. NK cells recognize and kill human glioblastoma cells with stem cell-like properties. J Immunol. 2009 Mar 15;182(6):3530-9.
- Sanchez-Correa B, Morgado S, Gayoso I, et al. Human NK cells in acute myeloid leukaemia patients: analysis of NK cell-activating receptors and their ligands. Cancer Immunol Immunother. 2011 Aug;60(8):1195-205.
- Imai K, Matsuyama S, Miyake S, Suga K, Nakachi K. Natural cytotoxic activity of peripheral-blood lymphocytes and cancer incidence: an 11-year follow-up study of a general population. Lancet. 2000 Nov 25;356(9244):1795-9.
- Solana R, Alonso MC, Peña J. Natural killer cells in healthy aging. Exp Gerontol. 1999 Jun;34(3):435-43.
- Beli E, Duriancik DM, Clinthorne JF, Lee T, Kim S, Gardner EM. Natural killer cell development and maturation in aged mice. Mech Ageing Dev. 2014 Jan;135:33-40.
- Chiu BC, Martin BE, Stolberg VR, Chensue SW. The host environment is responsible for aging-related functional NK cell deficiency. J Immunol. 2013 Nov 1;191(9):4688-98.
- Camous X, Pera A, Solana R, Larbi A. NK cells in healthy aging and age-associated diseases. J Biomed Biotechnol. 2012;2012:195956.
- Ihara T, Kamiya H, Starr SE, Arbeter AM, Lange B. Natural killing of varicella-zoster virus (VZV)-infected fibroblasts in normal children, children with VZV infections, and children with Hodgkin’s disease. Acta Paediatr Jpn. 1989 Oct;31(5):523-8.
- Guo H, Kumar P, Malarkannan S. Evasion of natural killer cells by influenza virus. J Leukoc Biol. 2011 Feb;89(2):189-94.
- Wilkinson GW, Tomasec P, Stanton RJ, et al. Modulation of natural killer cells by human cytomegalovirus. J Clin Virol. 2008 Mar;41(3):206-12.
- Simpson RJ. Aging, persistent viral infections, and immunosenescence: can exercise “make space”? Exerc Sport Sci Rev. 2011 Jan;39(1):23-33.
- Sampathkumar P, Drage LA, Martin DP. Herpes zoster (shingles) and postherpetic neuralgia. Mayo Clin Proc. 2009 Mar;84(3):274-80.
- Staras SA, Dollard SC, Radford KW, Flanders WD, Pass RF, Cannon MJ. Seroprevalence of cytomegalovirus infection in the United States, 1988-1994. Clin Infect Dis. 2006 Nov 1;43(9):1143-51.
- Savva GM, Pachnio A, Kaul B, et al. Cytomegalovirus infection is associated with increased mortality in the older population. Aging Cell. 2013 Jun;12(3):381-7.
- Smithey MJ, Li G, Venturi V, Davenport MP, Nikolich-Žugich J. Lifelong persistent viral infection alters the naive T cell pool, impairing CD8 T cell immunity in late life. J Immunol. 2012 Dec 1;189(11):5356-66.
- Weinberger B, Lazuardi L, Weiskirchner I, et al. Healthy aging and latent infection with CMV lead to distinct changes in CD8+ and CD4+ T-cell subsets in the elderly. Hum Immunol. 2007 Feb;68(2):86-90.
- Fletcher JM, Vukmanovic-Stejic M, Dunne PJ, et al. Cytomegalovirus-specific CD4+ T-cells in healthy carriers are continuously driven to replicative exhaustion. J Immunol. 2005 Dec 15;175(12):8218-25.
- Campos C, Pera A, Sanchez-Correa B, et al. Effect of age and CMV on NK cell subpopulations. Exp Gerontol. 2014 Jun;54:130-7.
- Pourgheysari B, Khan N, Best D, Bruton R, Nayak L, Moss PA. The cytomegalovirus-specific CD4+ T-cell response expands with age and markedly alters the CD4+ T-cell repertoire. J Virol. 2007 Jul;81(14):7759-65.
- Iannello A, Thompson TW, Ardolino M, Lowe SW, Raulet DH. p53-dependent chemokine production by senescent tumor cells supports NKG2D-dependent tumor elimination by natural killer cells. J Exp Med. 2013 Sep 23;210(10):2057-69.
- Smyth MJ, Wallace ME, Nutt SL, Yagita H, Godfrey DI, Hayakawa Y. Sequential activation of NKT-cells and NK cells provides effective innate immunotherapy of cancer. J Exp Med. 2005 Jun 20;201(12):1973-85.
- Raulet DH, Guerra N. Oncogenic stress sensed by the immune system: role of natural killer cell receptors. Nat Rev Immunol. 2009 Aug;9(8):568-80.
- Campos C, Pera A, Lopez-Fernandez I, Alonso C, Tarazona R, Solana R. Proinflammatory status influences NK cells subsets in the elderly. Immunol Lett. 2014 Jul 3.
- Thorén FB, Riise RE, Ousbäck J, et al. Human NK Cells induce neutrophil apoptosis via an NKp46- and Fas-dependent mechanism. J Immunol. 2012 Feb 15;188(4):1668-74.
- Iannello A, Raulet DH. Immunosurveillance of senescent cancer cells by natural killer cells. Oncoimmunology. 2014;3:e27616.
- Ferrando-Martínez S, Ruiz-Mateos E, Hernández A, et al. Age-related deregulation of naive T-cell homeostasis in elderly humans. Age (Dordr). 2011 Jun;33(2): 197-207.
- Kovaiou RD, Weiskirchner I, Keller M, Pfister G, Cioca DP, Grubeck-Loebenstein B. Age-related differences in phenotype and function of CD4+ T-cells are due to a phenotypic shift from naive to memory effector CD4+ T-cells. Int Immunol. 2005 Oct;17(10):1359-66.
- Wherry EJ, Blattman JN, Murali-Krishna K, van der Most R, Ahmed R. Viral persistence alters CD8 T-cell immunodominance and tissue distribution and results in distinct stages of functional impairment. J Virol. 2003 Apr;77(8):4911-27.
- Effros RB, Dagarag M, Spaulding C, Man J. The role of CD8+ T-cell replicative senescence in human aging. Immunol Rev. 2005 Jun;205:147-57.
- Scheller J, Ohnesorge N, Rose-John S. Interleukin-6 trans-signalling in chronic inflammation and cancer. Scand J Immunol. 2006 May;63(5):321-9.
- Montecino-Rodriguez E, Barent-Maoz B, Dorshkind K. Causes, consequences, and reversal of immune system aging. J Clin Invest. 2013;123(3):958-65.
- Wikby A, Maxson P, Olsson J, Johansson B, Ferguson FG. Changes in CD8 and CD4 lymphocyte subsets, T-cell proliferation responses and non-survival in the very old: the Swedish longitudinal OCTO-immune study. Mech Ageing Dev. 1998 May 15;102(2-3):187-98.
- Ferguson FG, Wikby A, Maxson P, Olsson J, Johansson B. Immune parameters in a longitudinal study of a very old population of Swedish people: a comparison between survivors and nonsurvivors. J Gerontol A Biol Sci Med Sci. 1995 Nov;50(6):B378-82.
- Kang I, Hong MS, Nolasco H, et al. Age-associated change in the frequency of memory CD4+ T-cells impairs long term CD4+ T-cell responses to influenza vaccine. J Immunol. 2004 Jul 1;173(1):673-81.
- Grubeck-Loebenstein B, Della Bella S, Iorio AM, Michel JP, Pawelec G, Solana R. Immunosenescence and vaccine failure in the elderly. Aging Clin Exp Res. 2009 Jun;21(3):201-9.
- Saurwein-Teissl M, Lung TL, Marx F, et al. Lack of antibody production following immunization in old age: association with CD8(+)CD28(-) T-cell clonal expansions and an imbalance in the production of Th1 and Th2 cytokines. J Immunol. 2002 Jun 1;168(11):5893-9.
- Wehrens EJ, Prakken BJ, van Wijk F. T-cells out of control--impaired immune regulation in the inflamed joint. Nat Rev Rheumatol. 2013 Jan;9(1):34-42.
- Ito TK, Yokoyama M, Yoshida Y, et al. A crucial role for CDC42 in senescence associated inflammation and atherosclerosis. PLoS One. 2014 Jul 24;9(7):e102186.
- Blankenberg S, Rupprecht HJ, Bickel C, Hafner G, Meyer J. The role of inflammation and infection in acute coronary syndrome. Herz. 2001 Apr;26 Suppl 1:9-18.
- Minamino T, Miyauchi H, Yoshida T, Ishida Y, Yoshida H, Komuro I. Endothelial cell senescence in human atherosclerosis: role of telomere in endothelial dysfunction. Circulation. 2002 Apr 2;105(13): 1541-4.
- Liang KP, Gabriel SE. Autoantibodies: innocent bystander or key player in immunosenescence and atherosclerosis? J Rheumatol. 2007 Jun;34(6):1203-7.
- Michel JJ, Turesson C, Lemster B, Atkins SR, Iclozan C, Bongartz T,et al. CD56-expressing T-cells that have features of senescence are expanded in rheumatoid arthritis. Arthritis Rheum. 2007 Jan;56(1):43-57.
- Schmidt D, Martens PB, Weyand CM, Goronzy JJ. The repertoire of CD4+ CD28- T-cells in rheumatoid arthritis. Mol Med. 1996 Sep;2(5):608-18.
- Schirmer M, Goldberger C, Würzner R, et al. Circulating cytotoxic CD8(+) CD28(-) T-cells in ankylosing spondylitis. Arthritis Res. 2002 4(1):71-6.
- Warrington KJ, Vallejo AN, Weyand CM, Goronzy JJ. CD28 loss in senescent CD4+ T-cells: reversal by interleukin-12 stimulation. Blood. 2003 May 1;101(9):3543-9.
- Moro-García MA, Echeverría A, Galán-Artímez MC, et al. Immunosenescence and inflammation characterize chronic heart failure patients with more advanced disease. Int J Cardiol. 2014 Jul 1;174(3):590-9.
- Voo KS, Peng G, Guo Z, et al. Functional characterization of EBV-encoded nuclear antigen 1-specific CD4+ helper and regulatory T-cells elicited by in vitro peptide stimulation. Cancer Res. 2005 Feb 15;65(4):1577-86.
- MacDonald TT. Suppressor T-cells, rebranded as regulatory T-cells, emerge from the wilderness bearing surface markers. Gut. 2002 Sep;51(3):311-2.
- Sojka DK, Huang YH, Fowell DJ. Mechanisms of regulatory T-cell suppression - a diverse arsenal for a moving target. Immunology. 2008 May;124(1):13-22.
- Available at: https://www.labcorp.com/wps/portal/!ut/p/c1/04. Accessed October 27, 2014.
- Müller L, Fülöp T, Pawelec G. Immunosenescence in vertebrates and invertebrates. Immun Ageing. 2013 Apr 2;10(1):12.
- Serrano-Villar S, Sainz T, Lee SA, et al. HIV-infected individuals with low CD4/CD8 ratio despite effective antiretroviral therapy exhibit altered T-cell subsets, heightened CD8+ T-cell activation, and increased risk of non-AIDS morbidity and mortality. PLoS Pathog. 2014 May 15;10(5):e1004078.
- Tancini G, Barni S, Rescaldani R, Fiorelli G, Vivani S, Lissoni P. Analysis of T helper and suppressor lymphocyte subsets in relation to the clinical stage of solid neoplasms. Oncology. 1990 47(5):381-4.
- Brivio F, Fumagalli L, Parolini D, et al. T-helper/T-regulator lymphocyte ratio as a new immunobiological index to quantify the anticancer immune status in cancer patients. In Vivo. 2008 Sep-Oct;22(5):647-50.
- Poggi A, Zocchi MR. Mechanisms of tumor escape: role of tumor microenvironment in inducing apoptosis of cytolytic effector cells. Arch Immunol Ther Exp (Warsz). 2006 Sep-Oct;54(5):323-33.
- Kim R, Emi M, Tanabe K. Cancer cell immune escape and tumor progression by exploitation of anti-inflammatory and pro-inflammatory responses. Cancer Biol Ther. 2005 Sep;4(9):924-33.
- Montes CL, Chapoval AI, Nelson J, et al. Tumor-induced senescent T-cells with suppressor function: a potential form of tumor immune evasion. Cancer Res. 2008 Feb 1;68(3):870-9.
- Petrini B, Wasserman J, Blomgren H, Rotstein S. Changes of blood T-cell subsets in patients receiving postoperative adjuvant chemotherapy for breast cancer. Eur J Cancer Clin Oncol. 1984 Dec;20(12):1485-7.
- Reizenstein P, Ogier C, Blomgren H, Petrini B, Wasserman J. Cells responsible for tumor surveillance in man: effects of radiotherapy, chemotherapy, and biologic response modifiers. Adv Immun Cancer Ther. 1985 1:1-28.
- Hegde S, Pahne J, Smola-Hess S. Novel immunosuppressive properties of interleukin-6 in dendritic cells: inhibition of NF-kappaB binding activity and CCR7 expression. FASEB J. 2004 Sep;18(12):1439-41.
- Mitchell WA, Meng I, Nicholson SA, Aspinall R. Thymic output, ageing and zinc. Biogerontology. 2006 Oct-Dec;7(5-6):461-70.
- May M, Holmes E, Rogers W, Poth M. Protection from glucocorticoid induced thymic involution by dehydroepiandrosterone. Life Sci. 1990 46(22):1627-31.
- Filipin Mdel V, Caetano LC, Brazão V, Santello FH, Toldo MP, do Prado JC Jr. DHEA and testosterone therapies in Trypanosoma cruzi-infected rats are associated with thymic changes. Res Vet Sci. 2010 Aug;89(1):98-103.
- Liu S, Ishikawa H, Li FJ, et al. Dehydroepiandrosterone can inhibit the proliferation of myeloma cells and the interleukin-6 production of bone marrow mononuclear cells from patients with myeloma. Cancer Res. 2005 Mar 15;65(6):2269-76.
- Araghi-Niknam M, Zhang Z, Jiang S, Call O, Eskelson CD, Watson RR. Cytokine dysregulation and increased oxidation is prevented by dehydroepiandrosterone in mice infected with murine leukemia retrovirus. Proc Soc Exp Biol Med. 1997 Dec;216(3):386-91.
- Jiang Y, Tu PF. Analysis of chemical constituents in Cistanche species. J Chromatogr A. 2009 Mar 13;1216(11):1970-9.
- Zhang K, Ma X, He W, et al. Extracts of cistanche deserticola can antagonize immunosenescence and extend life span in senescence-accelerated mouse prone 8 (SAM-P8) mice. Evid Based Complement Alternat Med. 2014;601383.
- Liang H, Yu F, Tong Z, Huang Z. Effect of Cistanches Herba aqueous extract on bone loss in ovariectomized rat. Int J Mol Sci. 2011 12(8):5060-9.
- Zeng JC, Fan YG, Liu JR, Zeng YR, Yi CZ, Yan L. Experimental study of directional differentiation of bone mesenchymal stem cells (BMSCs) to osteoblasts guided by serum containing cistanche deserticola. Zhongguo Gu Shang. 2010 Aug;23(8):606-8.
- Liang HD, Yu F, Tong ZH, Zhang HQ, Liang W. Cistanches Herba aqueous extract affecting serum BGP and TRAP and bone marrow Smad1 mRNA, Smad5 mRNA, TGF-β1 mRNA and TIEG1 mRNA expression levels in osteoporosis disease. Mol Biol Rep. 2013 Feb;40(2):757-63.
- van den Biggelaar AH, Huizinga TW, de Craen AJ, et al. Impaired innate immunity predicts frailty in old age. The Leiden 85-plus study. Exp Gerontol. 2004 Sep;39(9):1407-14.
- Yonei Y, Kitano T, Ogura1 M, et al. Effects of Health Food Containing Cistanche Deserticola Extract on QOL and Safety in Elderly: An Open Pilot Study of 12-week Oral Treatment. Anti-Aging Medicine. 2011 8(2):7-14.
- Iannello A, Raulet DH. Immune surveillance of unhealthy cells by natural killer cells. Cold Spring Harb Symp Quant Biol. 2013 78:249-57 .
- Krizhanovsky V, Xue W, Zender L, Yon M, Hernando E, Lowe SW. Implications of cellular senescence in tissue damage response, tumor suppression, and stem cell biology. Cold Spring Harb Symp Quant Biol. 2008 73:513-22.
- Clements ME, Chaber CJ, Ledbetter SR, Zuk A. Increased cellular senescence and vascular rarefaction exacerbate the progression of kidney fibrosis in aged mice following transient ischemic injury. PLoS One. 2013 Aug 5;8(8):e70464.
- Kumar M, Seeger W, Voswinckel R. Senescence-associated secretory phenotype and its possible role in chronic obstructive pulmonary disease. Am J Respir Cell Mol Biol. 2014 Sep;51(3):323-33.
- Bhat R, Crowe EP, Bitto A, et al. Astrocyte senescence as a component of Alzheimer’s disease. PLoS One. 2012 7(9):e45069.
- Boyd AR, Orihuela CJ. Dysregulated inflammation as a risk factor for pneumonia in the elderly. Aging Dis. 2011 Dec;2(6):487-500.
- Aspinall R, Andrew D. Thymic involution in aging. J Clin Immunol. 2000 Jul;20(4):250-6.
- Haase H, Rink L. The immune system and the impact of zinc during aging. Immun Ageing. 2009 Jun 12;6:9.
- Wong CP, Song Y, Elias VD, Magnusson KR, Ho E. Zinc supplementation increases zinc status and thymopoiesis in aged mice. J Nutr. 2009 Jul;139(7):1393-7.
- Dardenne M, Boukaiba N, Gagnerault MC, et al. Restoration of the thymus in aging mice by in vivo zinc supplementation. Clin Immunol Immunopathol. 1993 Feb;66(2):127-35.
- Available at: http://ods.od.nih.gov/factsheets/Zinc-HealthProfessional/. Accessed September 30, 2014.
- Ervin RB, Kennedy-Stephenson J. Mineral intakes of elderly adult supplement and non-supplement users in the third national health and nutrition examination survey. J Nutr. 2002 Nov;132(11):3422-7.
- Mocchegiani E, Romeo J, Malavolta M, et al. Zinc: dietary intake and impact of supplementation on immune function in elderly. Age (Dordr). 2013 Jun;35(3):839-60.
- Duchateau J, Delepesse G, Vrijens R, Collet H. Beneficial effects of oral zinc supplementation on the immune response of old people. Am J Med. 1981 May;70(5):1001-4.
- Prasad AS. Zinc: An antioxidant and anti-inflammatory agent: Role of zinc in degenerative disorders of aging. J Trace Elem Med Biol. 2014 Aug 12.
- Ali KH, Melillo AB, Leonard SM, et al. An open-label, randomized clinical trial to assess the immunomodulatory activity of a novel oligosaccharide compound in healthy adults. Functional Foods in Health and Disease. 2012 2(7):265-79.
- Daiwa Pharmaceutical. NK cell immunomodulatory function by modified arabinoxylan rice bran (MGN-3/Biobran) at low concentration (500 mg/day = 7 mg/kg/day). 2012. Supplier unpublished or internal study.
- Cholujova D, Jakubikova J, Czako B, et al. MGN-3 arabinoxylan rice bran modulates innate immunity in multiple myeloma patients. Cancer Immunol Immunother. 2013 Mar;62(3):437-45.
- Bang MH, Van Riep T, Thinh NT, et al. Arabinoxylan rice bran (MGN-3) enhances the effects of interventional therapies for the treatment of hepatocellular carcinoma: a three-year randomized clinical trial. Anticancer Res. 2010 Dec;30(12):5145-51.
- Ghoneum M. Immunostimulation and cancer prevention. The abstract of the 7th International Congress on anti-Aging & Biomedical Technologies Conference Proceedings Manual. 1999 Drew Univ., USA.
- Baker DJ, Wijshake T, Tchkonia T, et al. Clearance of p16Ink4a-positive senescenT-cells delays ageing-associated disorders. Nature. 2011 Nov 2;479(7372):232-6.
- Brockmeyer NH, Kreuzfelder E, Chalabi N, et al. The immunomodulatory potency of cimetidine in healthy volunteers. Int J Clin Pharmacol Ther Toxicol. 1989 Sep;27(9):458-62.
- Matsumoto S, Imaeda Y, Umemoto S, Kobayashi K, Suzuki H, Okamoto T. Cimetidine increases survival of colorectal cancer patients with high levels of sialyl Lewis-X and sialyl Lewis-A epitope expression on tumour cells. Br J Cancer. 2002 Jan 21;86(2):161-7.
- Kelly MD, King J, Cherian M, et al. Randomized trial of preoperative cimetidine in patients with colorectal carcinoma with quantitative assessment of tumor-associated lymphocytes. Cancer. 1999 Apr 15; 85(8):1658-63.
- Links M, Clingan PR, Phadke, et al. A randomized trial of cimetidine with 5-fluorouracil and folinic acid in metastatic colorectal cancer. Eur J Surg Oncol. 1995 Oct; 21(5):523-5.
- Kikuchi Y, Kizawa I, Oomori K, et al. Effects of cimetidine on interleukin-2 production by peripheral blood lymphocytes in advanced ovarian carcinoma. Eur J Cancer Clin Oncol. 1988 Jul;24(7):1185-90.
- Hayashi A, Kobayashi K, Imaeda Y, Matsumoto S. Cimetidine inhibits the adhesion of cancer cells with sialyl Lewis epitope onto the vascular endothelium. Gan To Kagaku Ryoho. 2003 Oct;30(11):1788-90.
- Natori T, Sata M, Nagai R, et al. Cimetidine inhibits angiogenesis and suppresses tumor growth. Biomed Pharmacother. 2005 Jan-Feb; 59(1-2):56-60.
- Novaro GM, Aronow HD, Mayer-Sabik E, Griffin BP. Plasma homocysteine and calcific aortic valve disease. Heart. 2004 Jul;90(7):802-3.
- Beulens JW, Bots ML, Atsma F, et al. High dietary menaquinone intake is associated with reduced coronary calcification. Atherosclerosis. 2009 Apr;203(2):489-93.
- Geleijnse JM, Vermeer C, Grobbee DE, et al. Dietary intake of menaquinone is associated with a reduced risk of coronary heart disease: the Rotterdam Study. J Nutr. 2004 Nov;134(11):3100-5.
- Fusaro M, Noale M, Viola V, et al. Vitamin K Italian (VIKI) Dialysis Study Investigators. Vitamin K, vertebral fractures, vascular calcifications, and mortality. J Bone Miner Res. 2012 Nov;27(11):2271-8.
- Theuwissen E, Smit E, Vermeer C. The role of vitamin K in soft-tissue calcification. Adv Nutr. 2012 Mar 1;3(2):166-73.
- Koos R, Mahnken AH, Mühlenbruch G, et al. Relation of oral anticoagulation to cardiac valvular and coronary calcium assessed by multislice spiral computed tomography. Am J Cardiol. 2005 Sep 15;96(6):747-9.
- Schurgers LJ, Dissel PE, Spronk HM, et al. Role of vitamin K and vitamin K-dependent proteins in vascular calcification. Z Kardiol. 2001; 90(Suppl):357-63.
- Shimizu T, Choudhry MA, Szalay L, et al. Salutary effects of androstenediol on cardiac function and splanchnic perfusion after trauma-hemorrhage. Am J Physiol Regul Integr Comp Physiol. 2004 Aug;287(2):R386-90.
- Haring R, Baumeister SE, Völzke H, et al. Prospective inverse associations of sex hormone concentrations in men with biomarkers of inflammation and oxidative stress. J Androl. 2012 Sep-Oct;33(5):944-50.
- Lichte P, Pfeifer R, Werner BE, et al. Dehydroepiandrosterone modulates the inflammatory response in a bilateral femoral shaft fracture model. Eur J Med Res. 2014 May 19;19:27.
Wellness
Specialists
1-800-226-2370 - This service is FREE
7:30 AM - 12 AM (ET) Mon-Fri | 9 AM - 12 AM (ET) Sat-Sun